

Combining affinity purification by ADP-ribose-binding macro domains with mass spectrometry to define the mammalian ADP-ribosyl proteome
- PMID: 19246377
- PMCID: PMC2657436
- DOI: 10.1073/pnas.0900066106
Combining affinity purification by ADP-ribose-binding macro domains with mass spectrometry to define the mammalian ADP-ribosyl proteome
Abstract
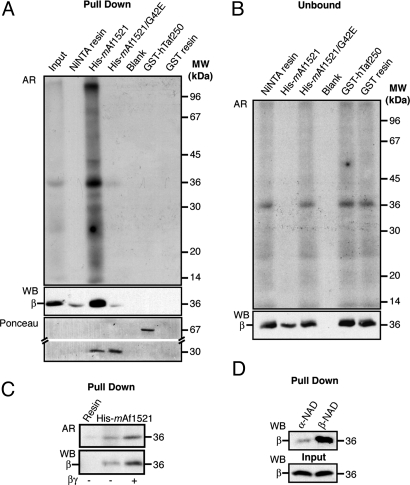
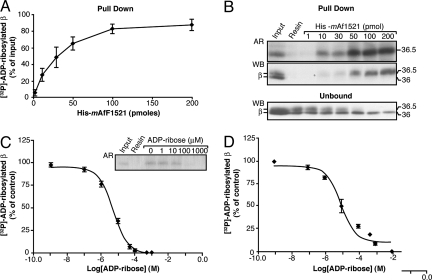
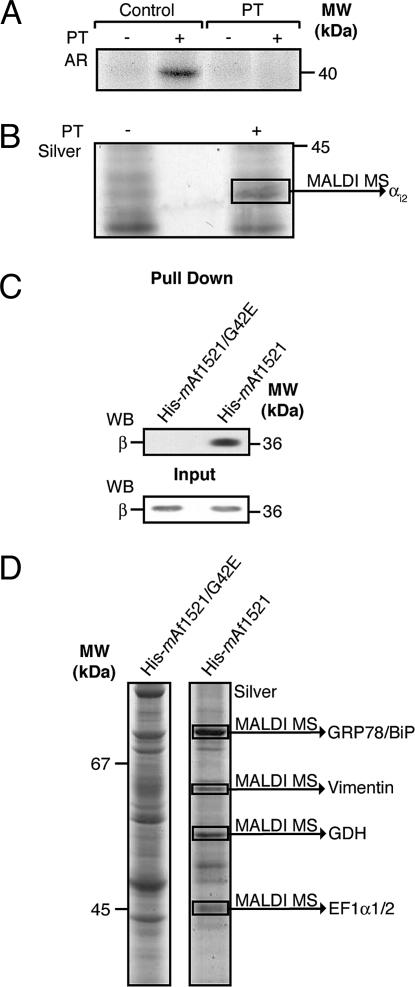
Mono-ADP-ribosylation is a reversible posttranslational modification that modulates the function of target proteins. The enzymes that catalyze this reaction in mammalian cells are either bacterial pathogenic toxins or endogenous cellular ADP-ribosyltransferases. For the latter, both the enzymes and their targets have largely remained elusive, mainly due to the lack of specific techniques to study this reaction. The recent discovery of the macro domain, a protein module that interacts selectively with ADP-ribose, prompted us to investigate whether this interaction can be extended to the identification of ADP-ribosylated proteins. Here, we report that macro domains can indeed be used as selective baits for high-affinity purification of mono-ADP-ribosylated proteins, which can then be identified by mass spectrometry. Using this approach, we have identified a series of cellular targets of ADP-ribosylation reactions catalyzed by cellular ADP-ribosyltransferases and toxins. These proteins include most of the known targets of ADP-ribosylation, indicating the validity of this method, and a large number of other proteins, which now need to be individually validated. This represents an important step toward the discovery of new ADP-ribosyltransferase targets and an understanding of the physiological role and the pharmacological potential of this protein modification.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
In Vitro Techniques for ADP-Ribosylated Substrate Identification.Methods Mol Biol. 2018;1813:25-40. doi: 10.1007/978-1-4939-8588-3_3. Methods Mol Biol. 2018. PMID: 30097859
-
Identification of ADP-ribosylated peptides and ADP-ribose acceptor sites.Front Biosci (Landmark Ed). 2014 Jun 1;19(7):1041-56. doi: 10.2741/4266. Front Biosci (Landmark Ed). 2014. PMID: 24896335 Review.
-
A role of intracellular mono-ADP-ribosylation in cancer biology.FEBS J. 2013 Aug;280(15):3551-62. doi: 10.1111/febs.12290. Epub 2013 May 10. FEBS J. 2013. PMID: 23590234 Review.
-
Generating Protein-Linked and Protein-Free Mono-, Oligo-, and Poly(ADP-Ribose) In Vitro.Methods Mol Biol. 2018;1813:91-108. doi: 10.1007/978-1-4939-8588-3_7. Methods Mol Biol. 2018. PMID: 30097863 Free PMC article.
-
Identification and analysis of ADP-ribosylated proteins.Curr Top Microbiol Immunol. 2015;384:33-50. doi: 10.1007/82_2014_424. Curr Top Microbiol Immunol. 2015. PMID: 25113886 Review.
Cited by
-
Macrodomain ADP-ribosylhydrolase and the pathogenesis of infectious diseases.PLoS Pathog. 2018 Mar 22;14(3):e1006864. doi: 10.1371/journal.ppat.1006864. eCollection 2018 Mar. PLoS Pathog. 2018. PMID: 29566066 Free PMC article. No abstract available.
-
Enhancing low-dose radiotherapy efficacy with PARP inhibitors via FBL-mediated oxidative stress response in colorectal cancer.Oncogene. 2025 Feb;44(4):228-240. doi: 10.1038/s41388-024-03207-w. Epub 2024 Nov 8. Oncogene. 2025. PMID: 39516657 Free PMC article.
-
PARP12-mediated ADP-ribosylation contributes to breast cancer cell fate by regulating AKT activation and DNA-damage response.Cell Mol Life Sci. 2025 Jan 23;82(1):58. doi: 10.1007/s00018-025-05586-z. Cell Mol Life Sci. 2025. PMID: 39847113 Free PMC article.
-
Overexpression of the WWE domain of RNF146 modulates poly-(ADP)-ribose dynamics at sites of DNA damage.DNA Repair (Amst). 2025 Jun;150:103845. doi: 10.1016/j.dnarep.2025.103845. Epub 2025 May 21. DNA Repair (Amst). 2025. PMID: 40403420 Free PMC article.
-
PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle.Cancers (Basel). 2022 Oct 24;14(21):5210. doi: 10.3390/cancers14215210. Cancers (Basel). 2022. PMID: 36358629 Free PMC article.
References
-
- Di Girolamo M, Dani N, Stilla A, Corda D. Physiological relevance of the endogenous mono(ADP-ribosyl)ation of cellular proteins. FEBS J. 2005;272:4565–4575. - PubMed
-
- Seman M, Adriouch S, Haag F, Koch-Nolte F. Ecto-ADP-ribosyltransferases (ARTs): Emerging actors in cell communication and signaling. Curr Med Chem. 2004;11:857–872. - PubMed
-
- Honjo T, Nishizuka Y, Hayaishi O. Diphtheria toxin-dependent adenosine diphosphate ribosylation of aminoacyl transferase II and inhibition of protein synthesis. J Biol Chem. 1968;243:3553–3555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
