

Exploring the roles of noise in the eukaryotic cell cycle
- PMID: 19246388
- PMCID: PMC2672517
- DOI: 10.1073/pnas.0810034106
Exploring the roles of noise in the eukaryotic cell cycle
Abstract
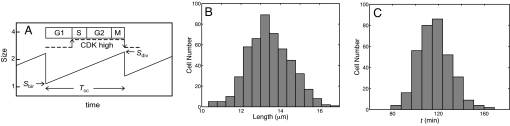
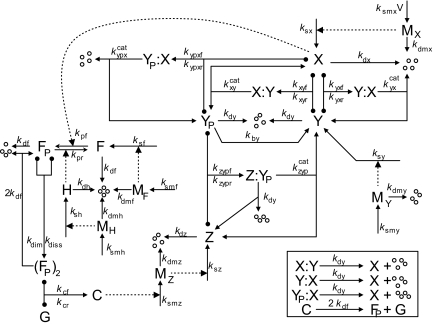
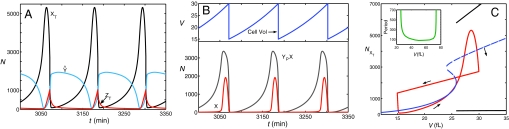
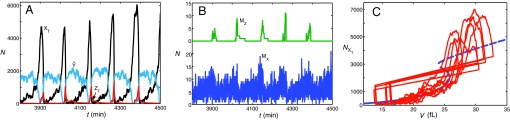
The DNA replication-division cycle of eukaryotic cells is controlled by a complex network of regulatory proteins, called cyclin-dependent kinases, and their activators and inhibitors. Although comprehensive and accurate deterministic models of the control system are available for yeast cells, reliable stochastic simulations have not been carried out because the full reaction network has yet to be expressed in terms of elementary reaction steps. As a first step in this direction, we present a simplified version of the control system that is suitable for exact stochastic simulation of intrinsic noise caused by molecular fluctuations and extrinsic noise because of unequal division. The model is consistent with many characteristic features of noisy cell cycle progression in yeast populations, including the observation that mRNAs are present in very low abundance (approximately 1 mRNA molecule per cell for each expressed gene). For the control system to operate reliably at such low mRNA levels, some specific mRNAs in our model must have very short half-lives (<1 min). If these mRNA molecules are longer-lived (perhaps 2 min), then the intrinsic noise in our simulations is too large, and there must be some additional noise suppression mechanisms at work in cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Tyson JJ. The coordination of cell growth and division: Intentional or incidental? Bioessays. 1985;2:72–77.
-
- Fantes PA. Control of cell size and cycle time in Schizosaccharomyces pombe. J Cell Sci. 1977;24:51–67. - PubMed
-
- Koch AL, Schaechter M. A model for statistics of the cell division process. J Gen Microbiol. 1962;29:435–454. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
