

The D4Z4 macrosatellite repeat acts as a CTCF and A-type lamins-dependent insulator in facio-scapulo-humeral dystrophy
- PMID: 19247430
- PMCID: PMC2639723
- DOI: 10.1371/journal.pgen.1000394
The D4Z4 macrosatellite repeat acts as a CTCF and A-type lamins-dependent insulator in facio-scapulo-humeral dystrophy
Abstract
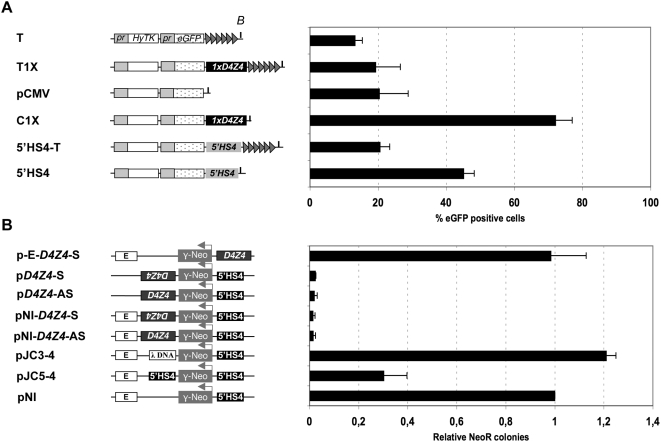
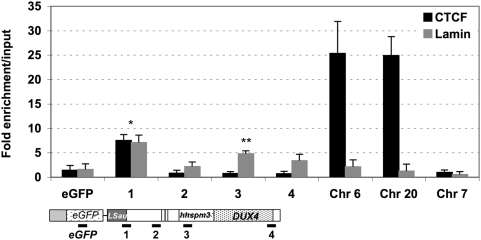
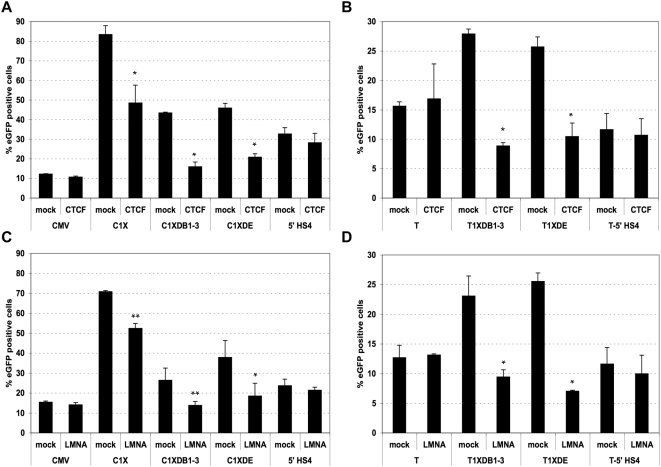
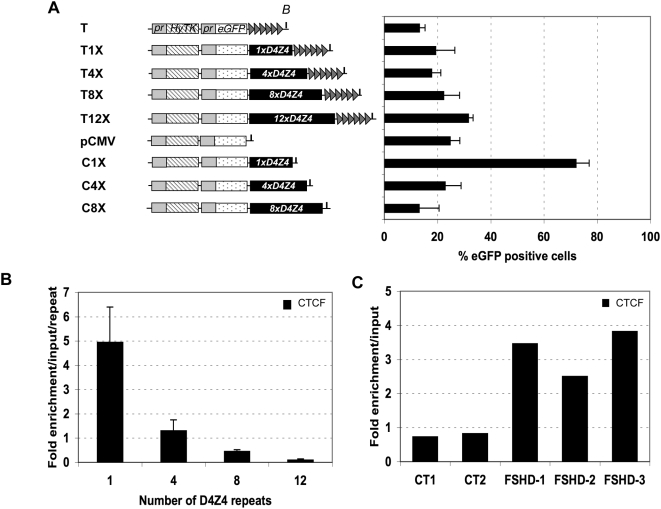
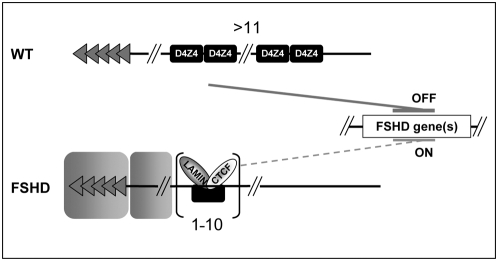
Both genetic and epigenetic alterations contribute to Facio-Scapulo-Humeral Dystrophy (FSHD), which is linked to the shortening of the array of D4Z4 repeats at the 4q35 locus. The consequence of this rearrangement remains enigmatic, but deletion of this 3.3-kb macrosatellite element might affect the expression of the FSHD-associated gene(s) through position effect mechanisms. We investigated this hypothesis by creating a large collection of constructs carrying 1 to >11 D4Z4 repeats integrated into the human genome, either at random sites or proximal to a telomere, mimicking thereby the organization of the 4q35 locus. We show that D4Z4 acts as an insulator that interferes with enhancer-promoter communication and protects transgenes from position effect. This last property depends on both CTCF and A-type Lamins. We further demonstrate that both anti-silencing activity of D4Z4 and CTCF binding are lost upon multimerization of the repeat in cells from FSHD patients compared to control myoblasts from healthy individuals, suggesting that FSHD corresponds to a gain-of-function of CTCF at the residual D4Z4 repeats. We propose that contraction of the D4Z4 array contributes to FSHD physio-pathology by acting as a CTCF-dependent insulator in patients.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Ottaviani A, Gilson E, Magdinier F. Telomeric position effect: From the yeast paradigm to human pathologies? Biochimie. 2008;90:93–107. - PubMed
-
- Wijmenga C, Hewitt JE, Sandkuijl LA, Clark LN, Wright TJ, et al. Chromosome 4q DNA rearrangements associated with facioscapulohumeral muscular dystrophy. Nat Genet. 1992;2:26–30. - PubMed
-
- Winokur ST, Bengtsson U, Feddersen J, Mathews KD, Weiffenbach B, et al. The DNA rearrangement associated with facioscapulohumeral muscular dystrophy involves a heterochromatin-associated repetitive element: implications for a role of chromatin structure in the pathogenesis of the disease. Chromosome Res. 1994;2:225–234. - PubMed
-
- van Deutekom JC, Wijmenga C, van Tienhoven EA, Gruter AM, Hewitt JE, et al. FSHD associated DNA rearrangements are due to deletions of integral copies of a 3.2 kb tandemly repeated unit. Hum Mol Genet. 1993;2:2037–2042. - PubMed
-
- Tawil R, Figlewicz DA, Griggs RC, Weiffenbach B. Facioscapulohumeral dystrophy: a distinct regional myopathy with a novel molecular pathogenesis. FSH Consortium. Ann Neurol. 1998;43:279–282. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
