

Molecular evolution of a viral non-coding sequence under the selective pressure of amiRNA-mediated silencing
- PMID: 19247440
- PMCID: PMC2642722
- DOI: 10.1371/journal.ppat.1000312
Molecular evolution of a viral non-coding sequence under the selective pressure of amiRNA-mediated silencing
Abstract
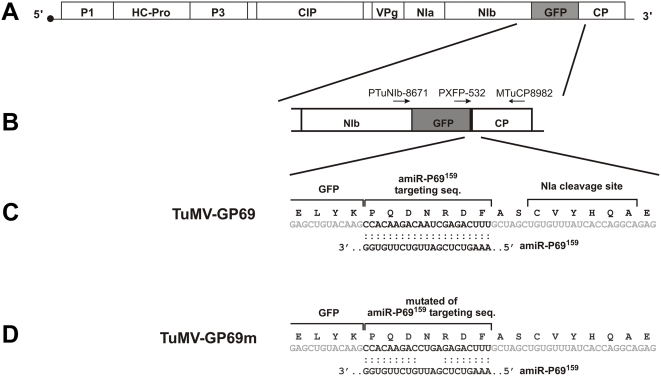
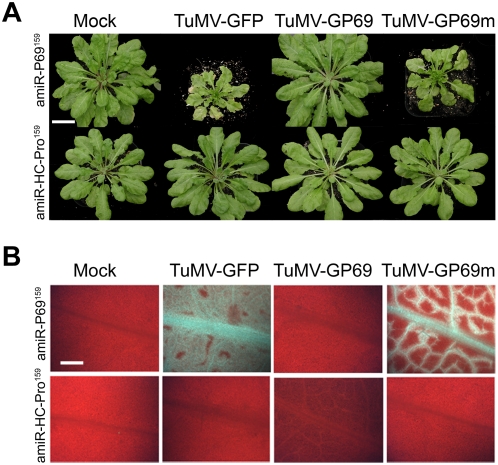
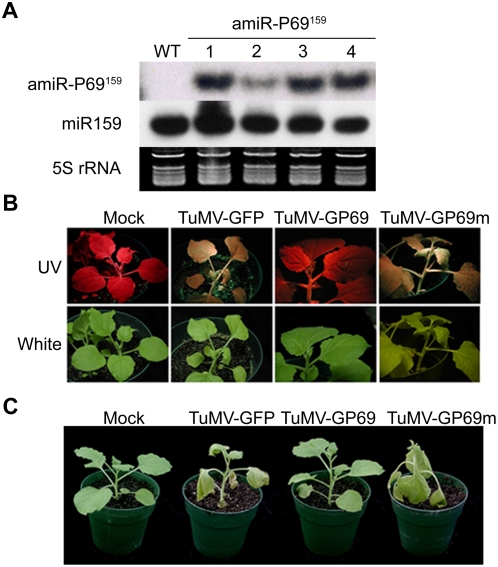
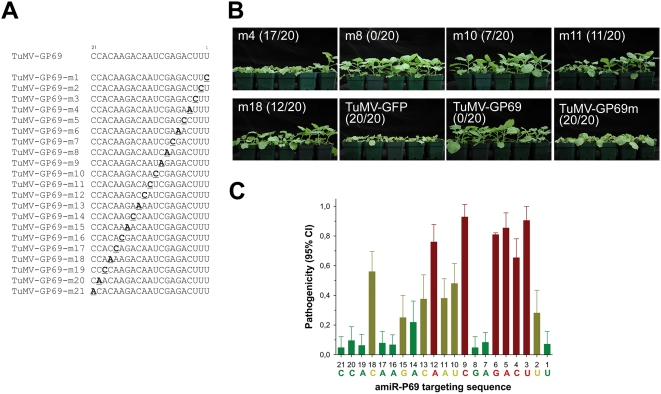
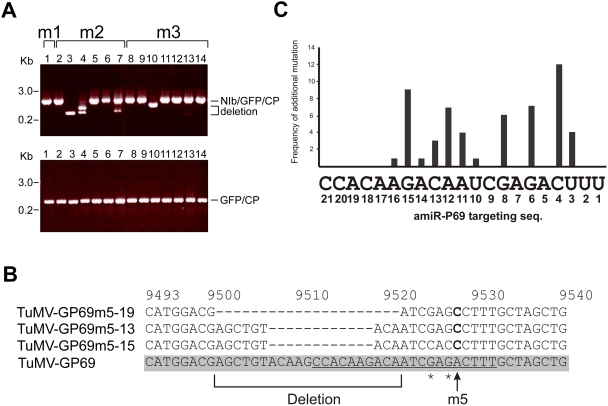
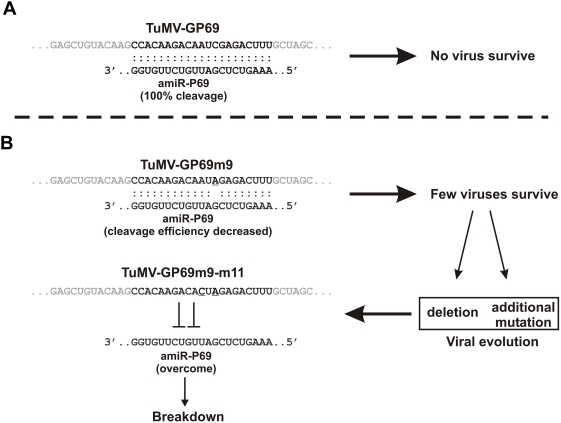
Plant microRNAs (miRNA) guide cleavage of target mRNAs by DICER-like proteins, thereby reducing mRNA abundance. Native precursor miRNAs can be redesigned to target RNAs of interest, and one application of such artificial microRNA (amiRNA) technology is to generate plants resistant to pathogenic viruses. Transgenic Arabidopsis plants expressing amiRNAs designed to target the genome of two unrelated viruses were resistant, in a highly specific manner, to the appropriate virus. Here, we pursued two different goals. First, we confirmed that the 21-nt target site of viral RNAs is both necessary and sufficient for resistance. Second, we studied the evolutionary stability of amiRNA-mediated resistance against a genetically plastic RNA virus, TuMV. To dissociate selective pressures acting upon protein function from those acting at the RNA level, we constructed a chimeric TuMV harboring a 21-nt, amiRNA target site in a non-essential region. In the first set of experiments designed to assess the likelihood of resistance breakdown, we explored the effect of single nucleotide mutation within the target 21-nt on the ability of mutant viruses to successfully infect amiRNA-expressing plants. We found non-equivalency of the target nucleotides, which can be divided into three categories depending on their impact in virus pathogenicity. In the second set of experiments, we investigated the evolution of the virus mutants in amiRNA-expressing plants. The most common outcome was the deletion of the target. However, when the 21-nt target was retained, viruses accumulated additional substitutions on it, further reducing the binding/cleavage ability of the amiRNA. The pattern of substitutions within the viral target was largely dominated by G to A and C to U transitions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Niu QW, Lin SS, Reyes JL, Chen KC, Wu HW, et al. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat Biotechnol. 2006;24:1420–1428. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
