

STIM1 clusters and activates CRAC channels via direct binding of a cytosolic domain to Orai1
- PMID: 19249086
- PMCID: PMC2670439
- DOI: 10.1016/j.cell.2009.02.014
STIM1 clusters and activates CRAC channels via direct binding of a cytosolic domain to Orai1
Abstract
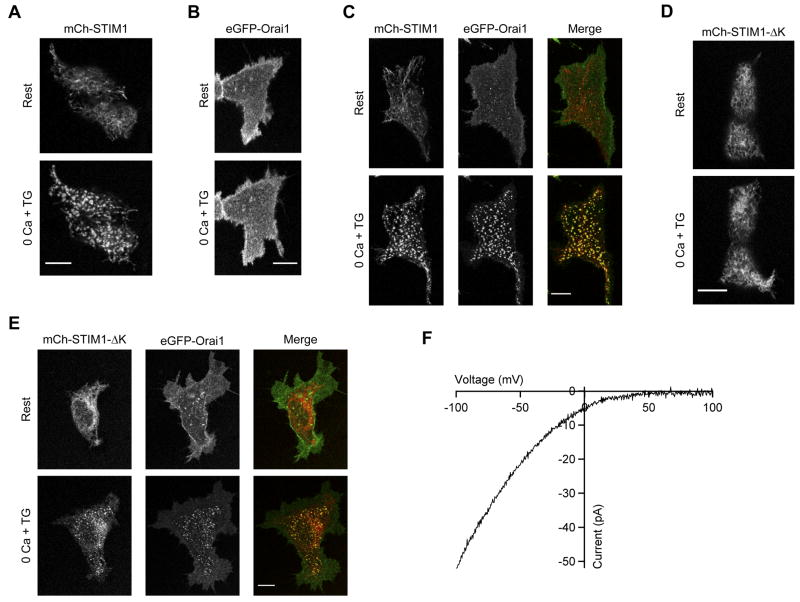
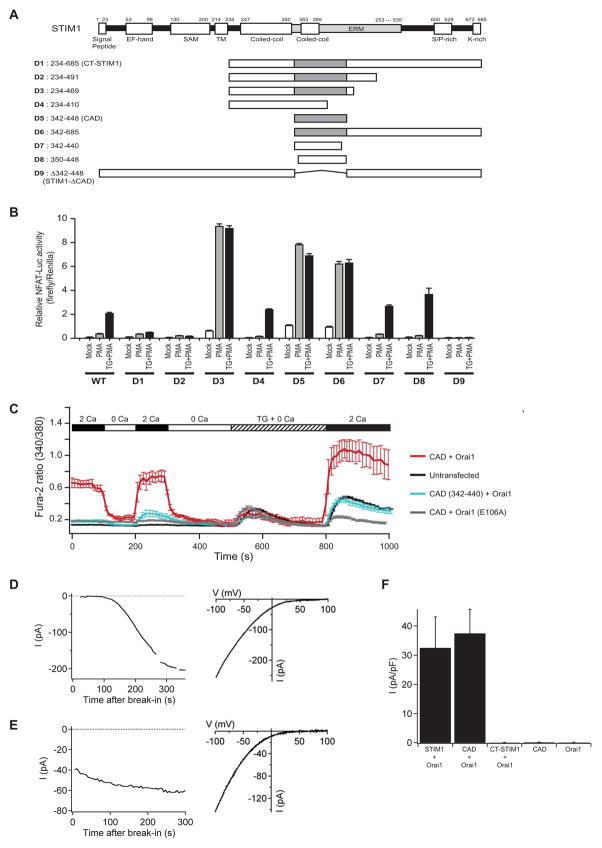
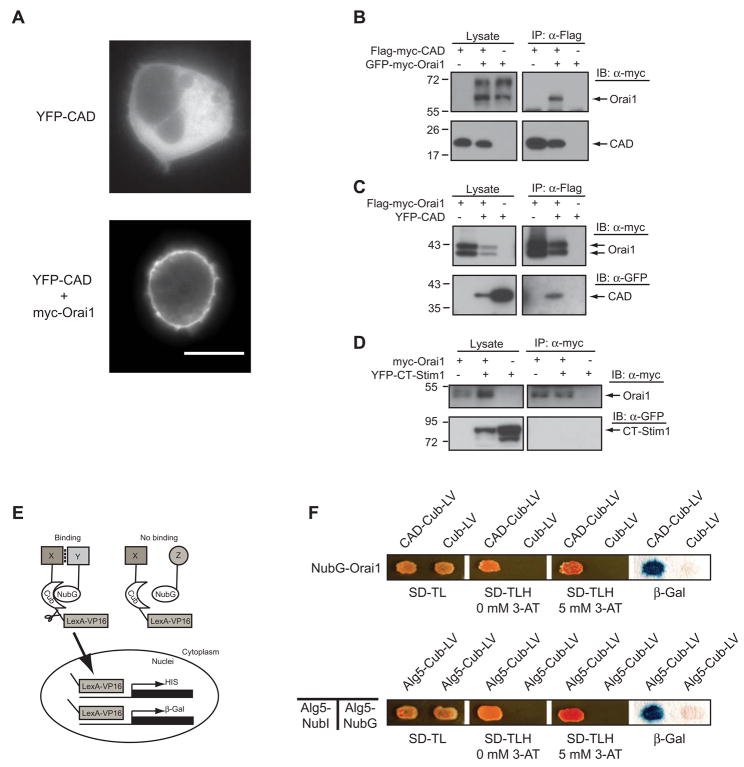
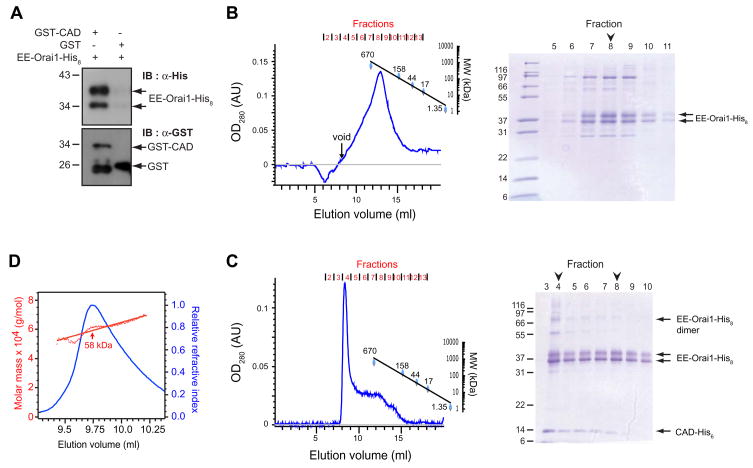
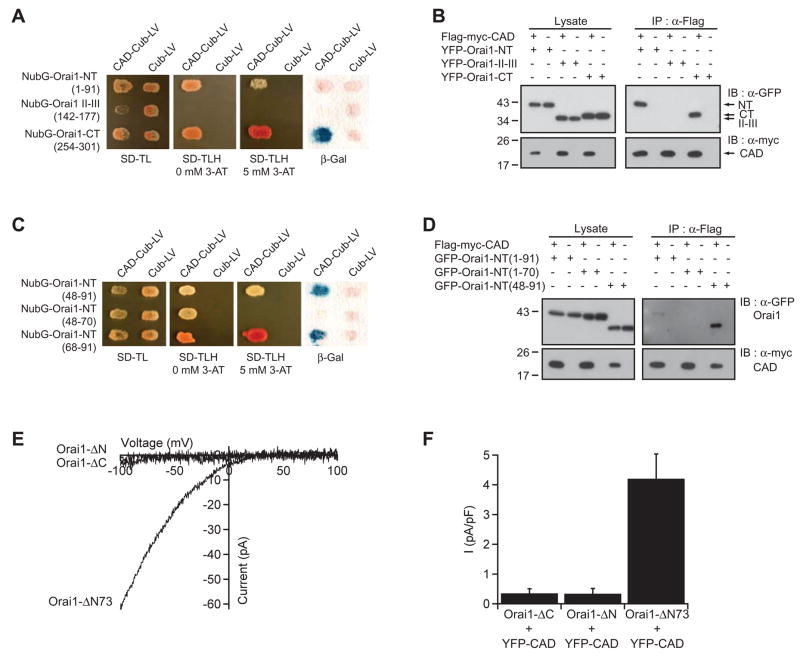
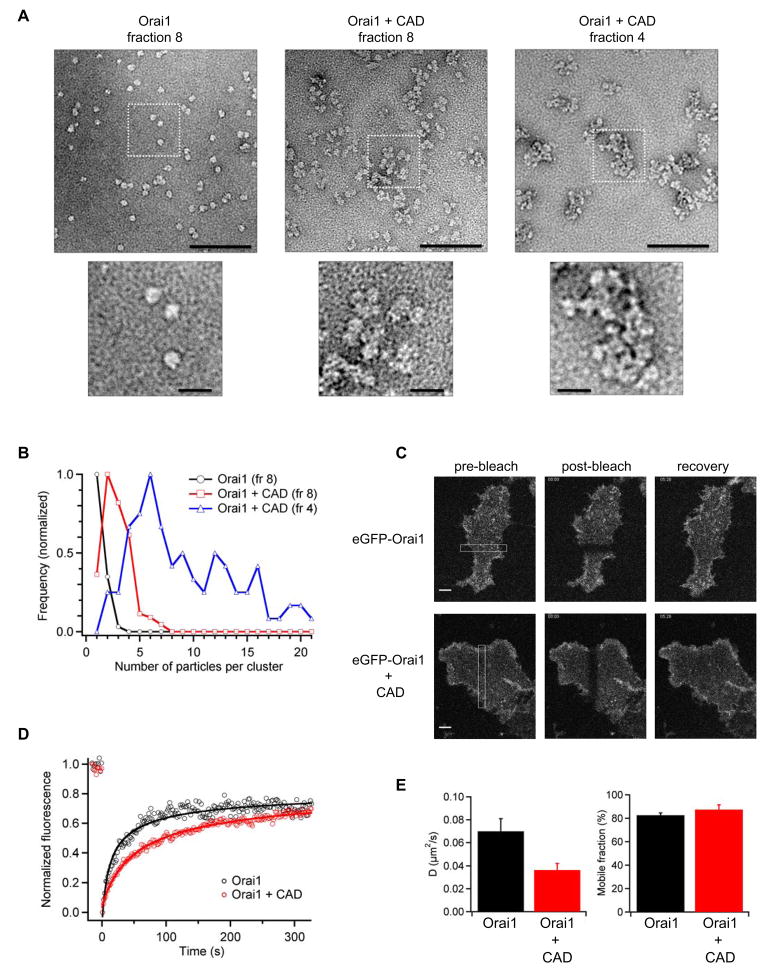
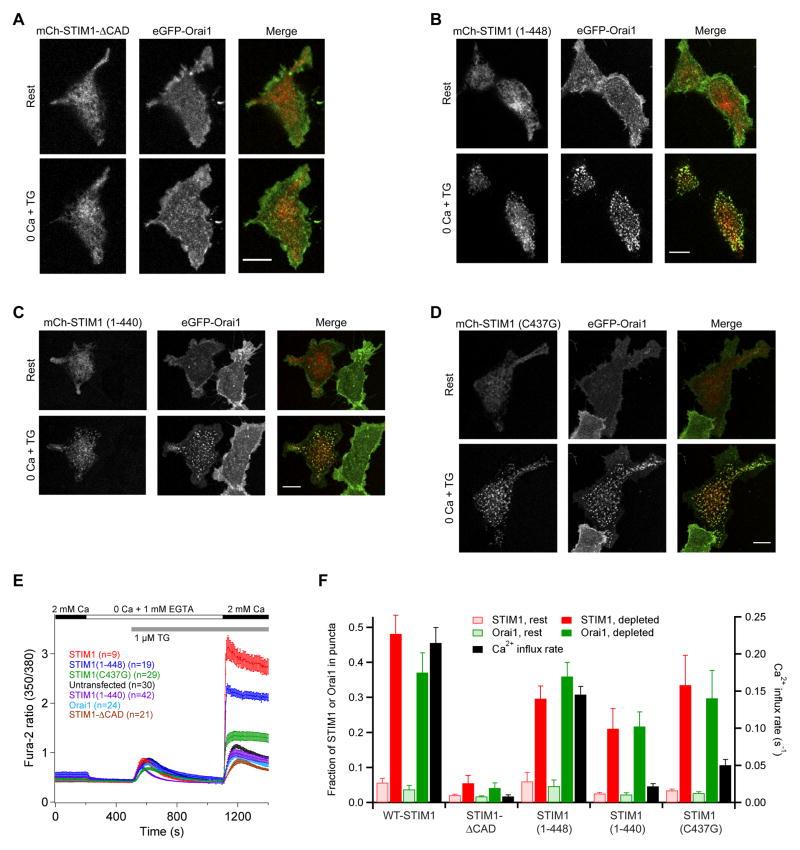
Store-operated Ca(2+) channels activated by the depletion of Ca(2+) from the endoplasmic reticulum (ER) are a major Ca(2+) entry pathway in nonexcitable cells and are essential for T cell activation and adaptive immunity. After store depletion, the ER Ca(2+) sensor STIM1 and the CRAC channel protein Orai1 redistribute to ER-plasma membrane (PM) junctions, but the fundamental issue of how STIM1 activates the CRAC channel at these sites is unresolved. Here, we identify a minimal, highly conserved 107-aa CRAC activation domain (CAD) of STIM1 that binds directly to the N and C termini of Orai1 to open the CRAC channel. Purified CAD forms a tetramer that clusters CRAC channels, but analysis of STIM1 mutants reveals that channel clustering is not sufficient for channel activation. These studies establish a molecular mechanism for store-operated Ca(2+) entry in which the direct binding of STIM1 to Orai1 drives the accumulation and the activation of CRAC channels at ER-PM junctions.
Figures
Comment in
-
A STIMulus Package puts orai calcium channels to work.Cell. 2009 Mar 6;136(5):814-6. doi: 10.1016/j.cell.2009.02.022. Cell. 2009. PMID: 19269360
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
