

Dscam mediates remodeling of glutamate receptors in Aplysia during de novo and learning-related synapse formation
- PMID: 19249274
- PMCID: PMC3442369
- DOI: 10.1016/j.neuron.2009.01.010
Dscam mediates remodeling of glutamate receptors in Aplysia during de novo and learning-related synapse formation
Abstract
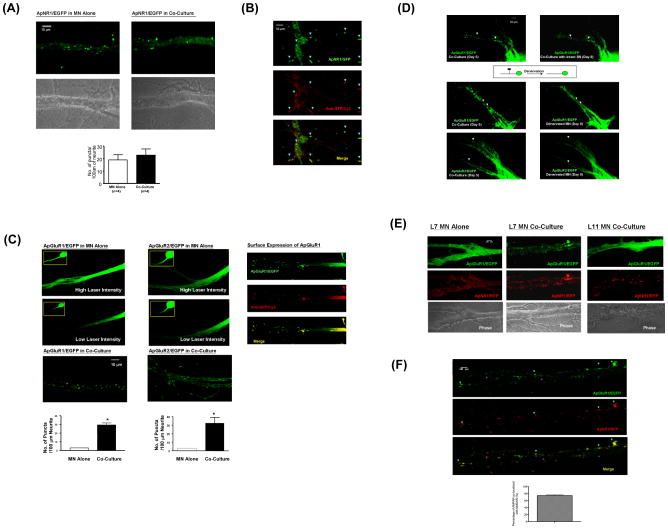
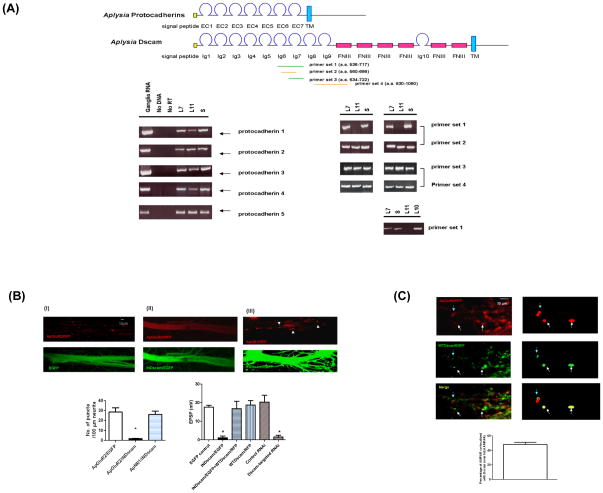
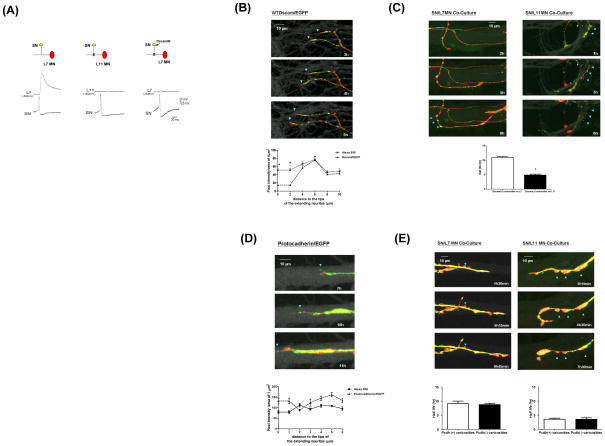
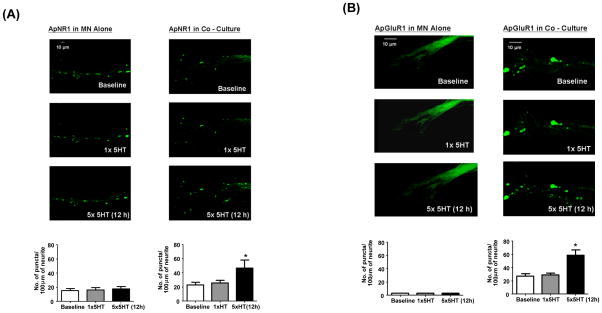
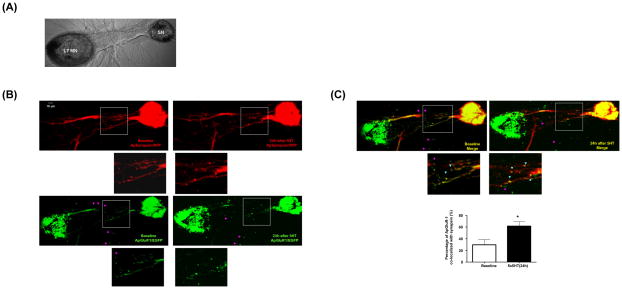
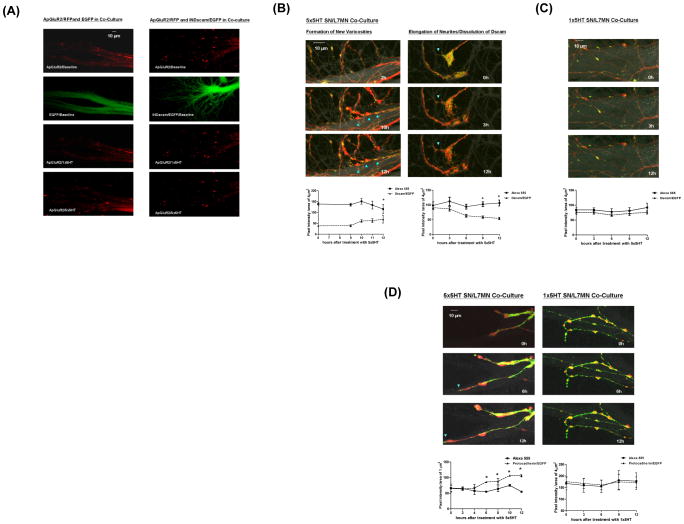
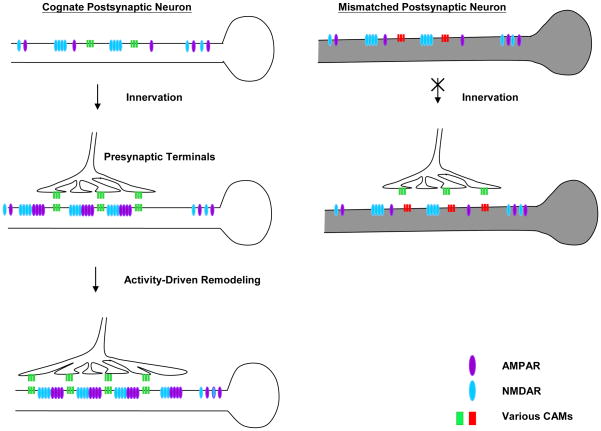
Transsynaptic interactions between neurons are essential during both developmental and learning-related synaptic growth. We have used Aplysia neuronal cultures to examine the contribution of transsynaptic signals in both types of synapse formation. We find that during de novo synaptogenesis, specific presynaptic innervation is required for the clustering of postsynaptic AMPA-like but not NMDA-like receptors. We further find that the cell adhesion molecule Dscam is involved in these transsynaptic interactions. Inhibition of Dscam either pre- or postsynaptically abolishes the emergence of synaptic transmission and the clustering of AMPA-like receptors. Remodeling of both AMPA-like and NMDA-like receptors also occurs during learning-related synapse formation and again requires the reactivation of Dscam-mediated transsynaptic interactions. Taken together, these findings suggest that learning-induced synapse formation recapitulates, at least in part, aspects of the mechanisms that govern de novo synaptogenesis.
Figures
Similar articles
-
Changes in functional glutamate receptors on a postsynaptic neuron accompany formation and maturation of an identified synapse.J Neurobiol. 1999 May;39(2):237-48. J Neurobiol. 1999. PMID: 10235678
-
Neurexin-neuroligin transsynaptic interaction mediates learning-related synaptic remodeling and long-term facilitation in aplysia.Neuron. 2011 May 12;70(3):468-81. doi: 10.1016/j.neuron.2011.03.020. Neuron. 2011. PMID: 21555073 Free PMC article.
-
Presynaptic and postsynaptic mechanisms of a novel form of homosynaptic potentiation at aplysia sensory-motor neuron synapses.J Neurosci. 2003 Aug 13;23(19):7288-97. doi: 10.1523/JNEUROSCI.23-19-07288.2003. J Neurosci. 2003. PMID: 12917362 Free PMC article.
-
The cellular mechanisms of learning in Aplysia: of blind men and elephants.Biol Bull. 2006 Jun;210(3):271-9. doi: 10.2307/4134563. Biol Bull. 2006. PMID: 16801500 Review.
-
Learning in Aplysia: looking at synaptic plasticity from both sides.Trends Neurosci. 2003 Dec;26(12):662-70. doi: 10.1016/j.tins.2003.09.014. Trends Neurosci. 2003. PMID: 14624850 Review.
Cited by
-
Reconsolidation of long-term memory in Aplysia.Curr Biol. 2012 Oct 9;22(19):1783-8. doi: 10.1016/j.cub.2012.07.038. Epub 2012 Aug 9. Curr Biol. 2012. PMID: 22885063 Free PMC article.
-
Identification of a serotonin receptor coupled to adenylyl cyclase involved in learning-related heterosynaptic facilitation in Aplysia.Proc Natl Acad Sci U S A. 2009 Aug 25;106(34):14634-9. doi: 10.1073/pnas.0907502106. Epub 2009 Aug 11. Proc Natl Acad Sci U S A. 2009. PMID: 19706550 Free PMC article.
-
DSCAM differentially modulates pre- and postsynaptic structural and functional central connectivity during visual system wiring.Neural Dev. 2018 Sep 15;13(1):22. doi: 10.1186/s13064-018-0118-5. Neural Dev. 2018. PMID: 30219101 Free PMC article.
-
Dysfunction of NMDA receptors in neuronal models of an autism spectrum disorder patient with a DSCAM mutation and in Dscam-knockout mice.Mol Psychiatry. 2021 Dec;26(12):7538-7549. doi: 10.1038/s41380-021-01216-9. Epub 2021 Jul 12. Mol Psychiatry. 2021. PMID: 34253863 Free PMC article.
-
Caffeine inhibits hypoxia-induced nuclear accumulation in HIF-1α and promotes neonatal neuronal survival.Exp Neurol. 2019 Jul;317:66-77. doi: 10.1016/j.expneurol.2019.01.014. Epub 2019 Feb 26. Exp Neurol. 2019. PMID: 30822423 Free PMC article.
References
-
- Agarwala KL, Ganesh S, Amano K, Suzuki T, Yamakawa K. DSCAM, a highly conserved gene in mammals, expressed in differentiating mouse brain. Biochem Biophys Res Commun. 2001;281:697–705. - PubMed
-
- Bailey CH, Alberini C, Ghirardi M, Kandel ER. Molecular and structural changes underlying long-term memory storage in Aplysia. Adv Second Messenger Phosphoprotein Res. 1994;29:529–544. - PubMed
-
- Bailey CH, Kandel ER. Structural changes accompanying memory storage. Annu Rev Physiol. 1993;55:397–426. - PubMed
-
- Barlow GM, Micales B, Chen XN, Lyons GE, Korenberg JR. Mammalian DSCAMs: roles in the development of the spinal cord, cortex, and cerebellum? Biochem Biophys Res Commun. 2002;293:881–891. - PubMed
-
- Barlow GM, Micales B, Lyons GE, Korenberg JR. Down syndrome cell adhesion molecule is conserved in mouse and highly expressed in the adult mouse brain. Cytogenet Cell Genet. 2001;94:155–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
