

A presynaptic homeostatic signaling system composed of the Eph receptor, ephexin, Cdc42, and CaV2.1 calcium channels
- PMID: 19249276
- PMCID: PMC2699049
- DOI: 10.1016/j.neuron.2008.12.028
A presynaptic homeostatic signaling system composed of the Eph receptor, ephexin, Cdc42, and CaV2.1 calcium channels
Abstract
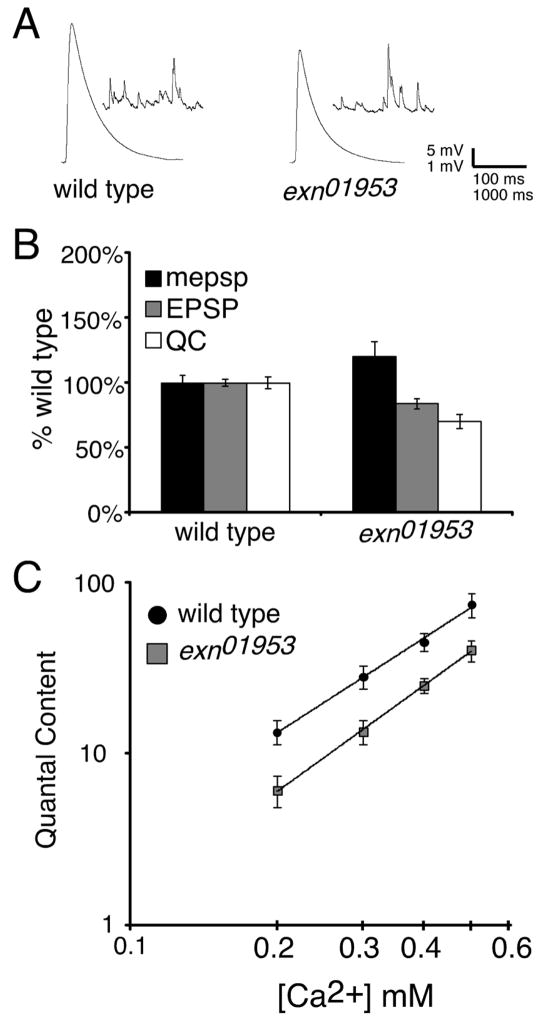
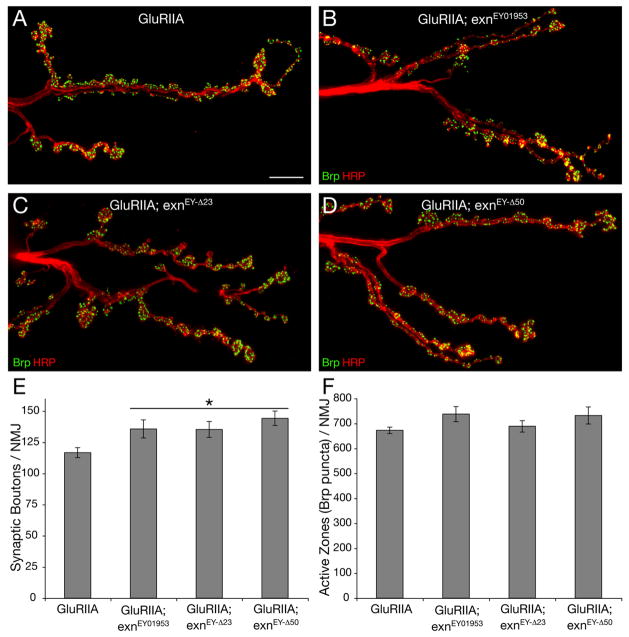
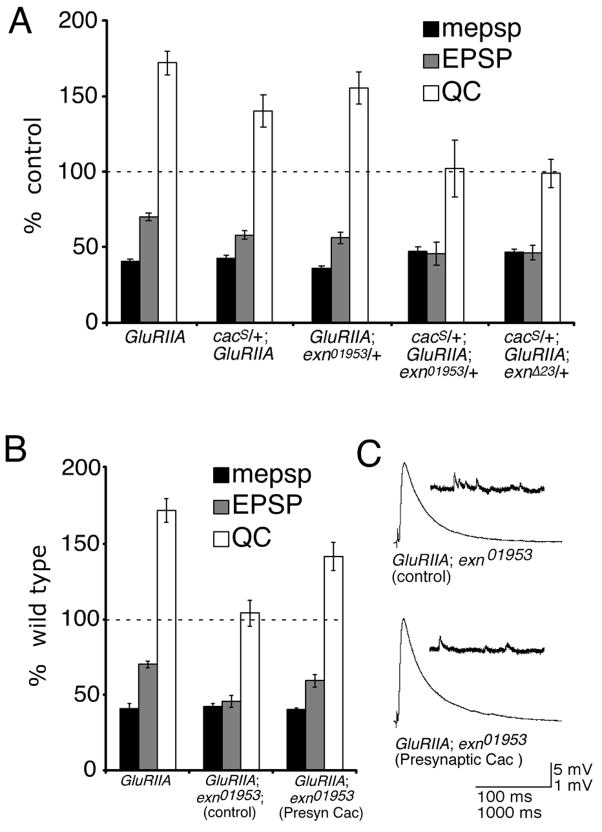
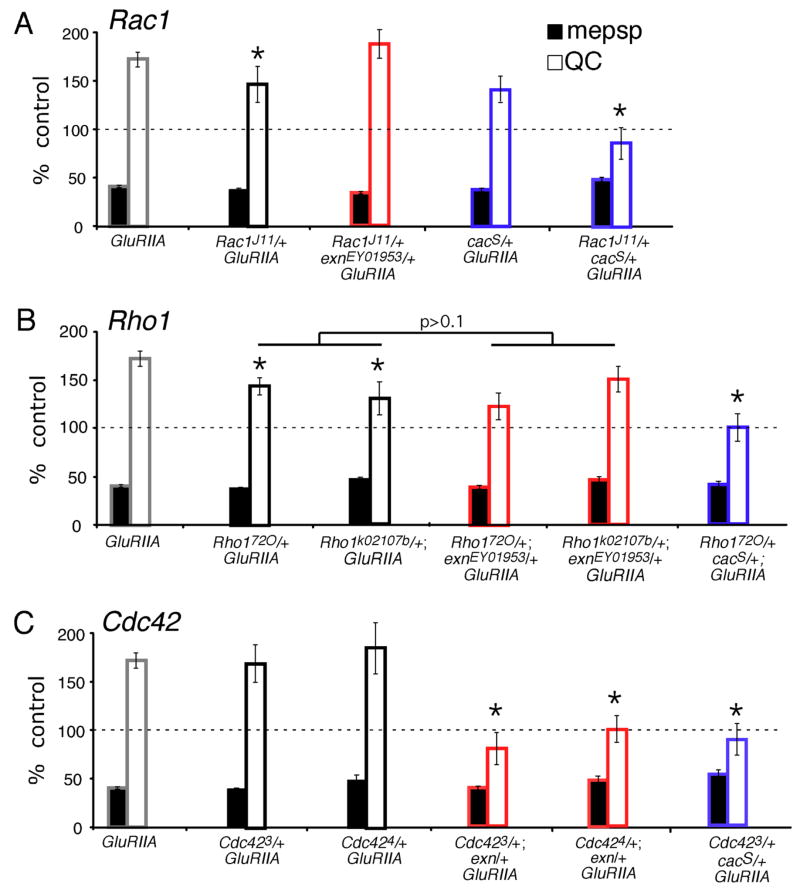
The molecular mechanisms underlying the homeostatic modulation of presynaptic neurotransmitter release remain largely unknown. In a screen, we isolated mutations in Drosophila ephexin (Rho-type guanine nucleotide exchange factor) that disrupt the homeostatic enhancement of presynaptic release following impairment of postsynaptic glutamate receptor function at the Drosophila neuromuscular junction. We show that Ephexin is sufficient presynaptically for synaptic homeostasis and localizes in puncta throughout the nerve terminal. However, ephexin mutations do not alter other aspects of neuromuscular development, including morphology or active zone number. We then show that, during synaptic homeostasis, Ephexin functions primarily with Cdc42 in a signaling system that converges upon the presynaptic CaV2.1 calcium channel. Finally, we show that Ephexin binds the Drosophila Eph receptor (Eph) and Eph mutants disrupt synaptic homeostasis. Based on these data, we propose that Ephexin/Cdc42 couples synaptic Eph signaling to the modulation of presynaptic CaV2.1 channels during the homeostatic enhancement of presynaptic release.
Figures
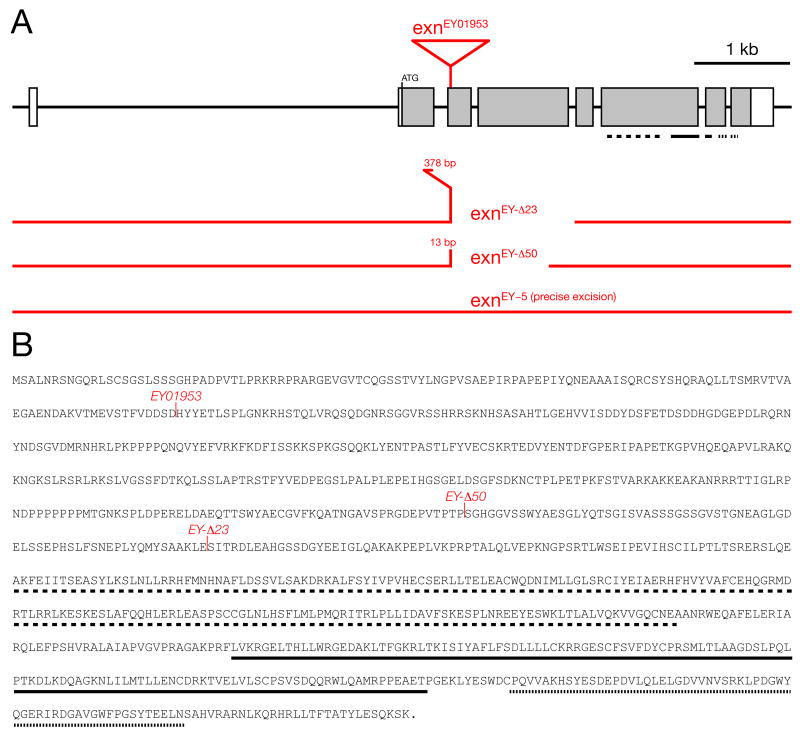
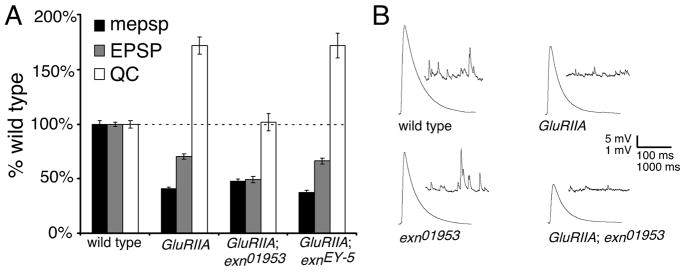
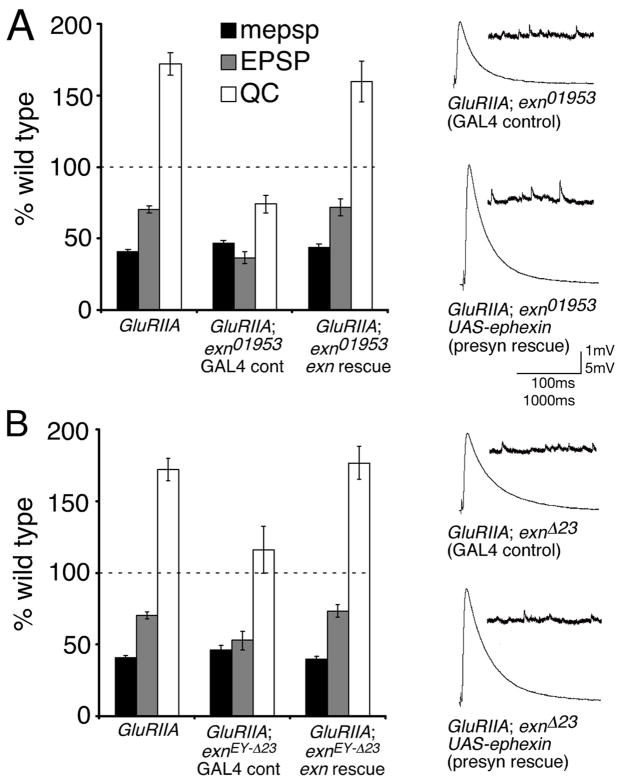
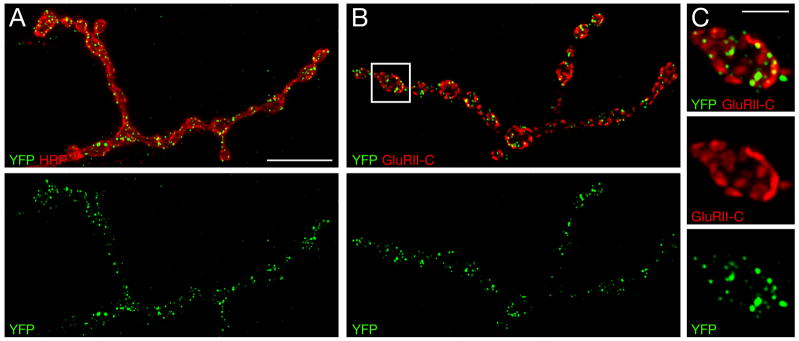
References
-
- Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhaes TR, Goodman CS. wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron. 2002;33:545–558. - PubMed
-
- Burrone J, Murthy VN. Synaptic gain control and homeostasis. Curr Opin Neurobiol. 2003;13:560–567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
