

E2-RING expansion of the NEDD8 cascade confers specificity to cullin modification
- PMID: 19250909
- PMCID: PMC2725360
- DOI: 10.1016/j.molcel.2009.01.011
E2-RING expansion of the NEDD8 cascade confers specificity to cullin modification
Abstract
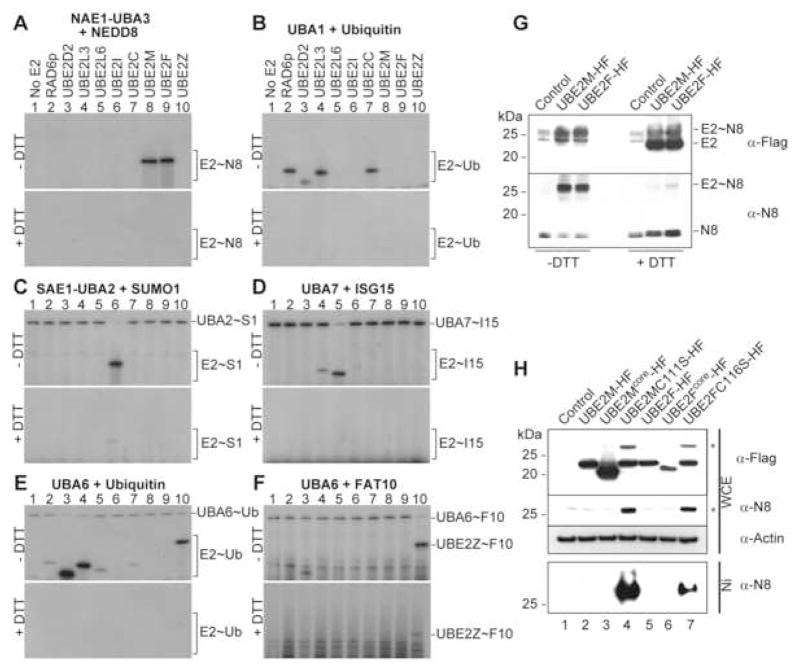
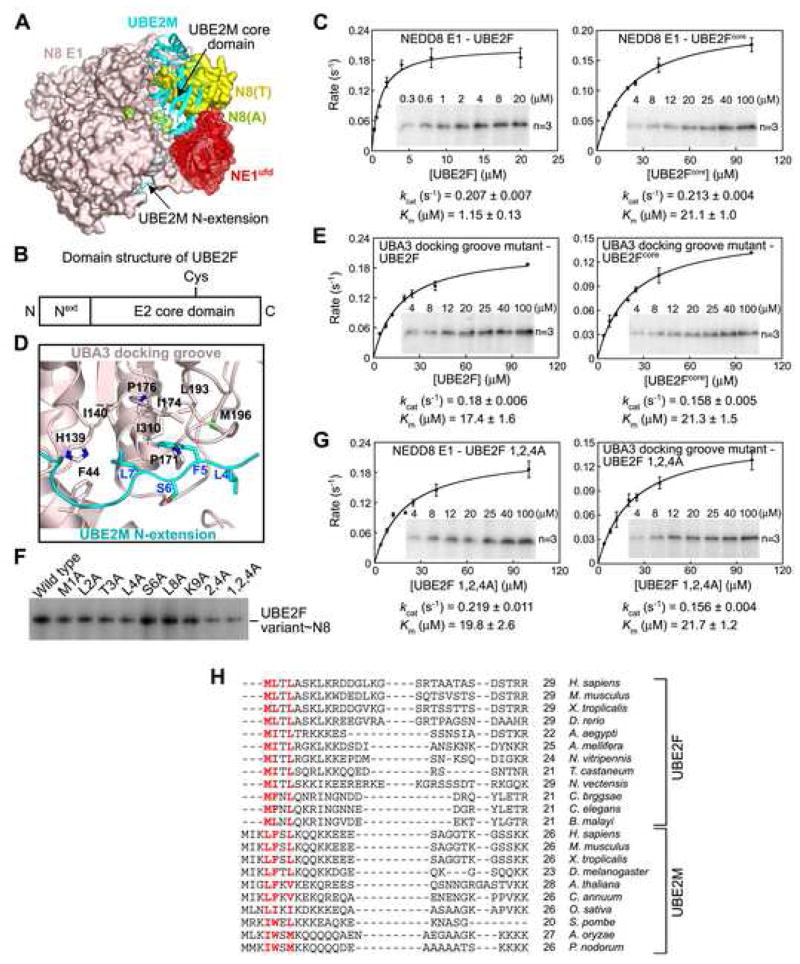
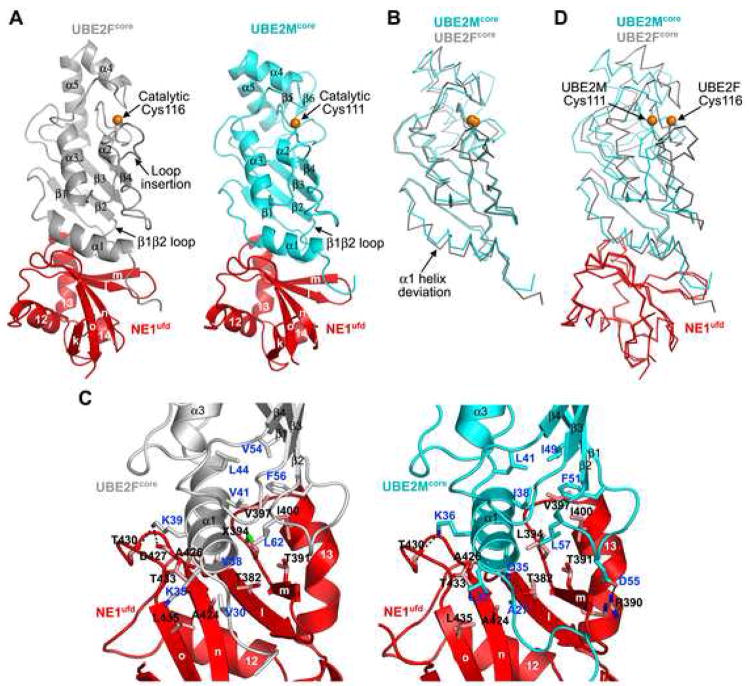
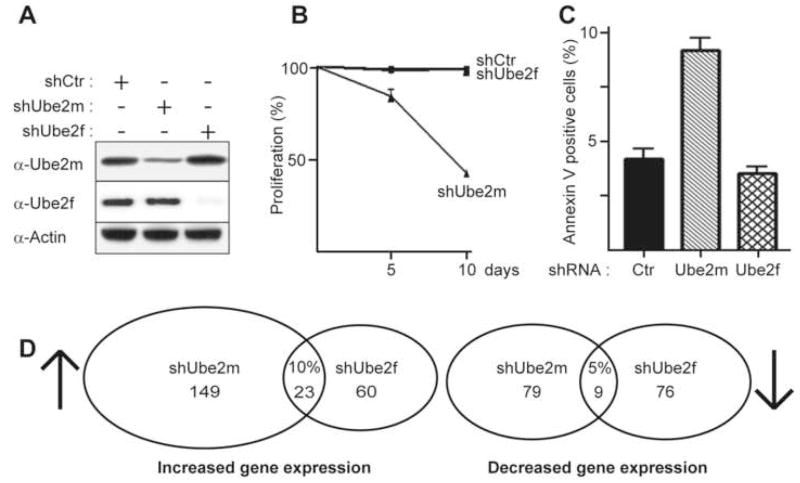
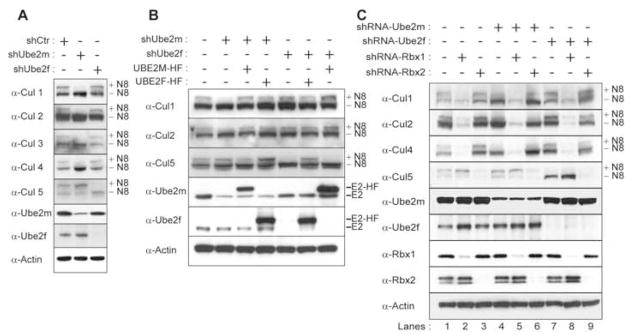
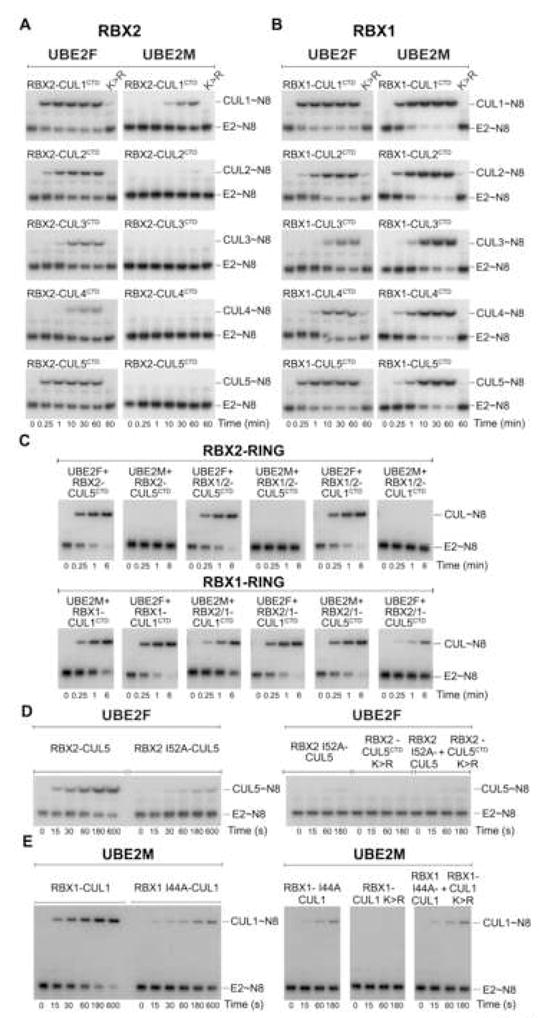
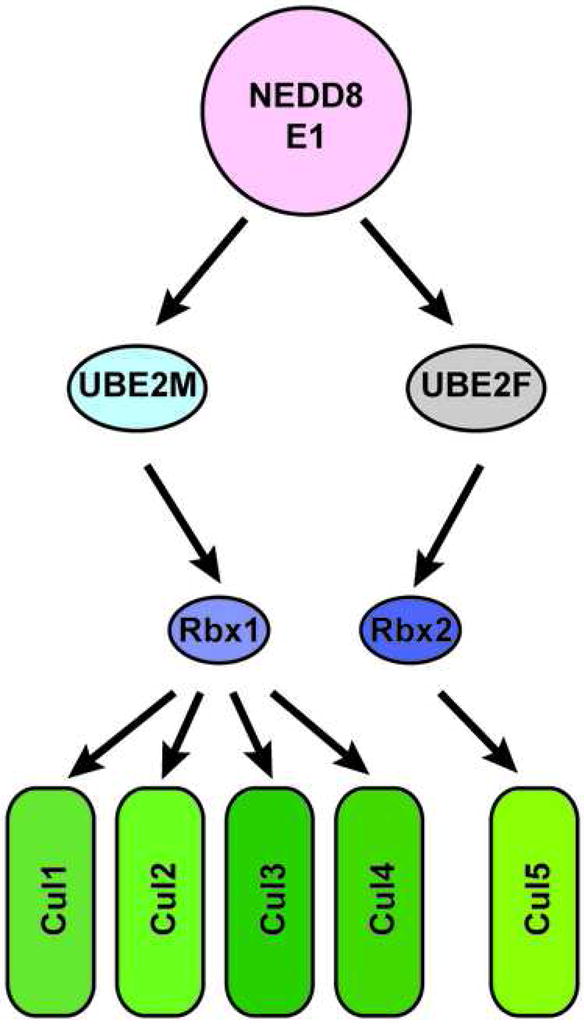
Ubiquitin and ubiquitin-like proteins (UBLs) are directed to targets by cascades of E1, E2, and E3 enzymes. The largest ubiquitin E3 subclass consists of cullin-RING ligases (CRLs), which contain one each of several cullins (CUL1, -2, -3, -4, or -5) and RING proteins (RBX1 or -2). CRLs are activated by ligation of the UBL NEDD8 to a conserved cullin lysine. How is cullin NEDD8ylation specificity established? Here we report that, like UBE2M (also known as UBC12), the previously uncharacterized E2 UBE2F is a NEDD8-conjugating enzyme in vitro and in vivo. Biochemical and structural analyses indicate how plasticity of hydrophobic E1-E2 interactions and E1 conformational flexibility allow one E1 to charge multiple E2s. The E2s have distinct functions, with UBE2M/RBX1 and UBE2F/RBX2 displaying different target cullin specificities. Together, these studies reveal the molecular basis for and functional importance of hierarchical expansion of the NEDD8 conjugation system in establishing selective CRL activation.
Figures
References
-
- The CCP4 suite: programs for protein crystallography. Acta crystallographica. 1994;50:760–763. - PubMed
-
- Amir RE, Iwai K, Ciechanover A. The NEDD8 pathway is essential for SCF(beta -TrCP)-mediated ubiquitination and processing of the NF-kappa B precursor p105. The Journal of biological chemistry. 2002;277:23253–23259. - PubMed
-
- Cardozo T, Pagano M. The SCF ubiquitin ligase: insights into a molecular machine. Nature reviews. 2004;5:739–751. - PubMed
-
- Chiu YH, Sun Q, Chen ZJ. E1-L2 activates both ubiquitin and FAT10. Molecular cell. 2007;27:1014–1023. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
