

Caenorhabditits elegans LRK-1 and PINK-1 act antagonistically in stress response and neurite outgrowth
- PMID: 19251702
- PMCID: PMC2713553
- DOI: 10.1074/jbc.M808255200
Caenorhabditits elegans LRK-1 and PINK-1 act antagonistically in stress response and neurite outgrowth
Abstract
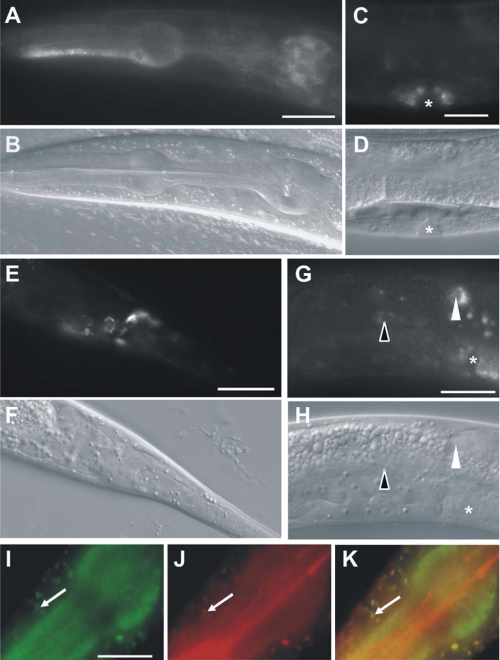
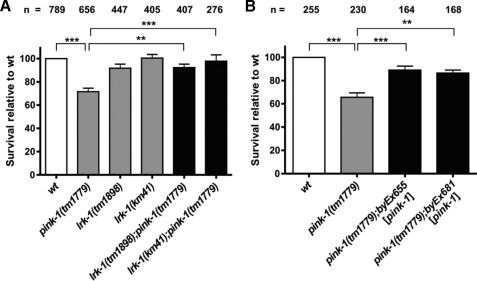
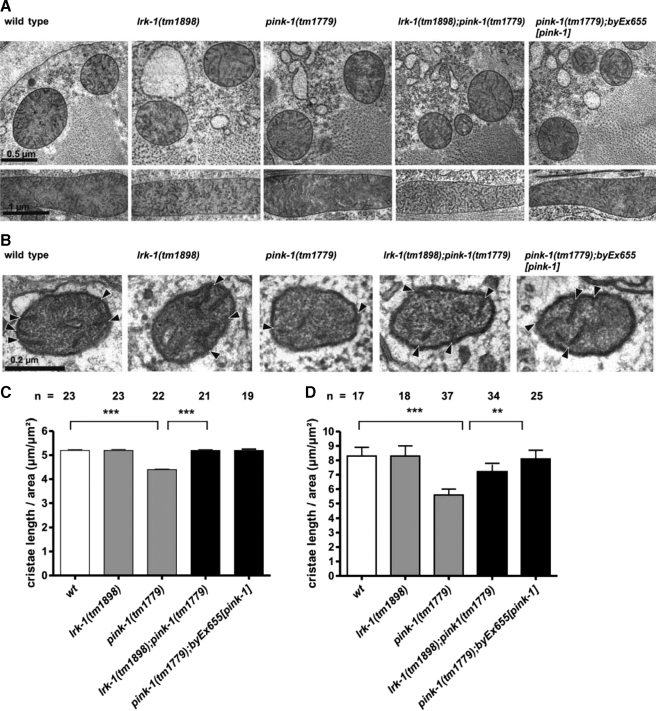
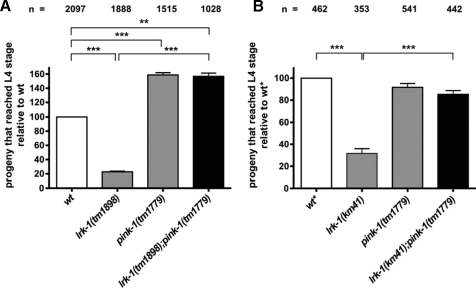
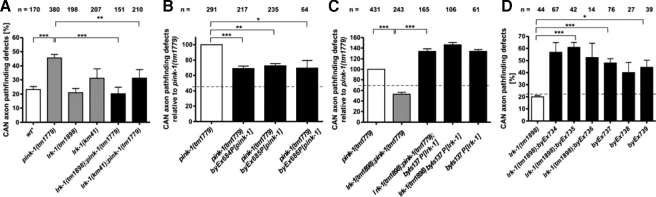
Mutations in two genes encoding the putative kinases LRRK2 and PINK1 have been associated with inherited variants of Parkinson disease. The physiological role of both proteins is not known at present, but studies in model organisms have linked their mutants to distinct aspects of mitochondrial dysfunction, increased vulnerability to oxidative and endoplasmic reticulum stress, and intracellular protein sorting. Here, we show that a mutation in the Caenorhabditits elegans homologue of the PTEN-induced kinase pink-1 gene resulted in reduced mitochondrial cristae length and increased paraquat sensitivity of the nematode. Moreover, the mutants also displayed defects in axonal outgrowth of a pair of canal-associated neurons. We demonstrate that in the absence of lrk-1, the C. elegans homologue of human LRRK2, all phenotypic aspects of pink-1 loss-of-function mutants were suppressed. Conversely, the hypersensitivity of lrk-1 mutant animals to the endoplasmic reticulum stressor tunicamycin was reduced in a pink-1 mutant background. These results provide the first evidence of an antagonistic role of PINK-1 and LRK-1. Due to the similarity of the C. elegans proteins to human LRRK2 and PINK1, we suggest a common role of both factors in cellular functions including stress response and regulation of neurite outgrowth. This study might help to link pink-1/PINK1 and lrk-1/LRRK2 function to the pathological processes resulting from Parkinson disease-related mutants in both genes, the first manifestations of which are cytoskeletal defects in affected neurons.
Figures
References
-
- Valente E. M., Abou-Sleiman P. M., Caputo V., Muqit M. M., Harvey K., Gispert S., Ali Z., Del T. D., Bentivoglio A. R., Healy D. G., Albanese A., Nussbaum R., Gonzalez-Maldonado R., Deller T., Salvi S., Cortelli P., Gilks W. P., Latchman D. S., Harvey R. J., Dallapiccola B., Auburger G., Wood N. W. ( 2004) Science 304, 1158– 1160 - PubMed
-
- Park J., Lee S. B., Lee S., Kim Y., Song S., Kim S., Bae E., Kim J., Shong M., Kim J. M., Chung J. ( 2006) Nature 441, 1157– 1161 - PubMed
-
- Clark I. E., Dodson M. W., Jiang C., Cao J. H., Huh J. R., Seol J. H., Yoo S. J., Hay B. A., Guo M. ( 2006) Nature 441, 1162– 1166 - PubMed
-
- Silvestri L., Caputo V., Bellacchio E., Atorino L., Dallapiccola B., Valente E. M., Casari G. ( 2005) Hum. Mol. Genet 14, 3477– 3492 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
