

GRAS proteins form a DNA binding complex to induce gene expression during nodulation signaling in Medicago truncatula
- PMID: 19252081
- PMCID: PMC2660633
- DOI: 10.1105/tpc.108.064501
GRAS proteins form a DNA binding complex to induce gene expression during nodulation signaling in Medicago truncatula
Abstract
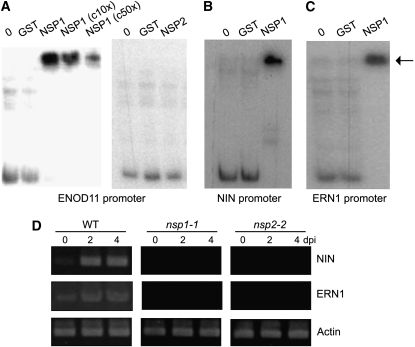
The symbiotic association of legumes with rhizobia involves bacterially derived Nod factor, which is sufficient to activate the formation of nodules on the roots of the host plant. Perception of Nod factor by root hair cells induces calcium oscillations that are a component of the Nod factor signal transduction pathway. Perception of the calcium oscillations is a function of a calcium- and calmodulin-dependent protein kinase, and this activates nodulation gene expression via two GRAS domain transcriptional regulators, Nodulation Signaling Pathway1 (NSP1) and NSP2, and an ERF transcription factor required for nodulation. Here, we show that NSP1 and NSP2 form a complex that is associated with the promoters of early nodulin genes. We show that NSP1 binds directly to ENOD promoters through the novel cis-element AATTT. While NSP1 shows direct binding to the ENOD11 promoter in vitro, this association in vivo requires NSP2. The NSP1-NSP2 association with the ENOD11 promoter is enhanced following Nod factor elicitation. Mutations in the domain of NSP2 responsible for its interaction with NSP1 highlight the significance of the NSP1-NSP2 heteropolymer for nodulation signaling. Our work reveals direct binding of a GRAS protein complex to DNA and highlights the importance of the NSP1-NSP2 complex for efficient nodulation in the model legume Medicago truncatula.
Figures






Comment in
-
Nodulation signaling in legumes depends on an NSP1-NSP2 complex.Plant Cell. 2009 Feb;21(2):367. doi: 10.1105/tpc.109.210214. Epub 2009 Feb 27. Plant Cell. 2009. PMID: 19252077 Free PMC article. No abstract available.
References
-
- Ane, J.M., et al. (2004). Medicago truncatula DMI1 required for bacterial and fungal symbioses in legumes. Science 303 1364–1367. - PubMed
-
- Boisson-Dernier, A., Andriankaja, A., Chabaud, M., Niebel, A., Journet, E.P., Barker, D.G., and de Carvalho-Niebel, F. (2005). MtENOD11 gene activation during rhizobial infection and mycorrhizal arbuscule development requires a common AT-rich-containing regulatory sequence. Mol. Plant Microbe Interact. 18 1269–1276. - PubMed
-
- Boisson-Dernier, A., Chabaud, M., Garcia, F., Becard, G., Rosenberg, C., and Barker, D.G. (2001). Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol. Plant Microbe Interact. 14 695–700. - PubMed
-
- Bolle, C. (2004). The role of GRAS proteins in plant signal transduction and development. Planta 218 683–692. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
