

Gene expression divergence in yeast is coupled to evolution of DNA-encoded nucleosome organization
- PMID: 19252487
- PMCID: PMC2744203
- DOI: 10.1038/ng.324
Gene expression divergence in yeast is coupled to evolution of DNA-encoded nucleosome organization
Abstract
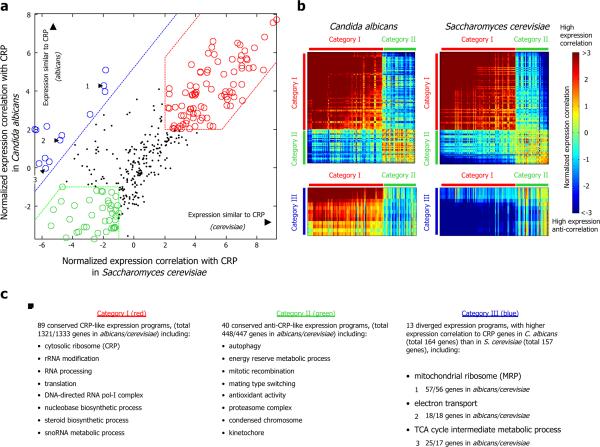
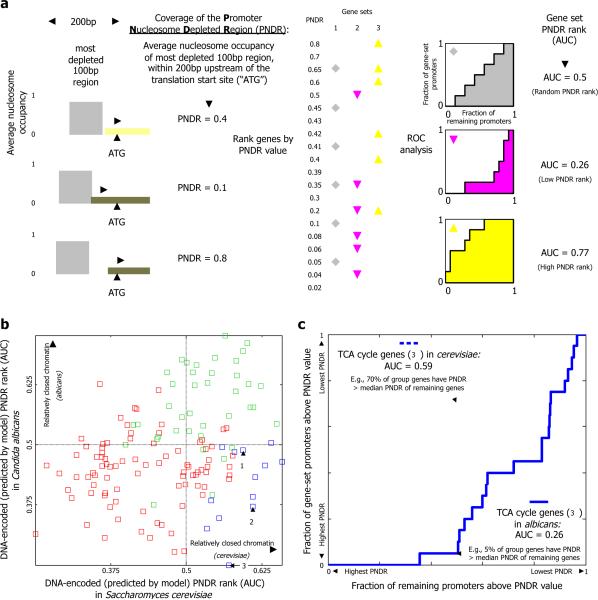
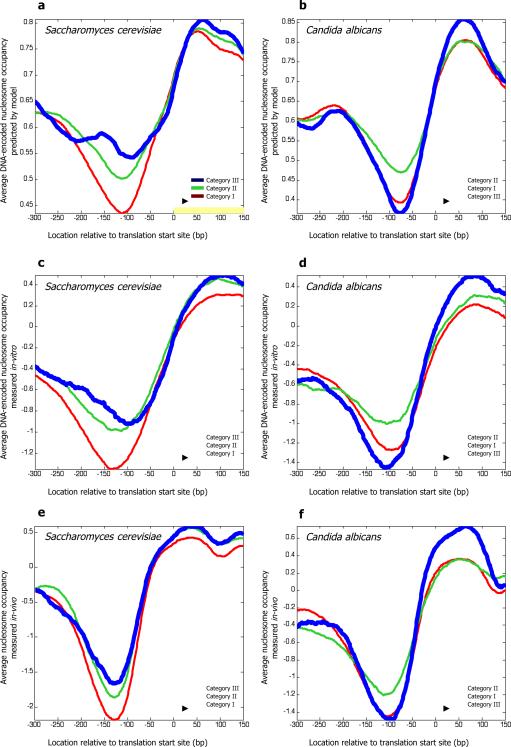
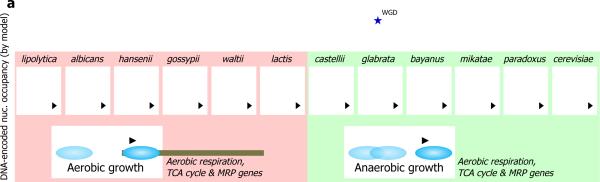
Eukaryotic transcription occurs within a chromatin environment, whose organization has an important regulatory function and is partly encoded in cis by the DNA sequence itself. Here, we examine whether evolutionary changes in gene expression are linked to changes in the DNA-encoded nucleosome organization of promoters. We find that in aerobic yeast species, where cellular respiration genes are active under typical growth conditions, the promoter sequences of these genes encode a relatively open (nucleosome-depleted) chromatin organization. This nucleosome-depleted organization requires only DNA sequence information, is independent of any cofactors and of transcription, and is a general property of growth-related genes. In contrast, in anaerobic yeast species, where cellular respiration genes are relatively inactive under typical growth conditions, respiration gene promoters encode relatively closed (nucleosome-occupied) chromatin organizations. Our results suggest a previously unidentified genetic mechanism underlying phenotypic diversity, consisting of DNA sequence changes that directly alter the DNA-encoded nucleosome organization of promoters.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
