

Potential role of TBC1D4 in enhanced post-exercise insulin action in human skeletal muscle
- PMID: 19252894
- PMCID: PMC3627047
- DOI: 10.1007/s00125-009-1294-y
Potential role of TBC1D4 in enhanced post-exercise insulin action in human skeletal muscle
Abstract
Aims/hypothesis: TBC1 domain family, member 4 (TBC1D4; also known as AS160) is a cellular signalling intermediate to glucose transport regulated by insulin-dependent and -independent mechanisms. Skeletal muscle insulin sensitivity is increased after acute exercise by an unknown mechanism that does not involve modulation at proximal insulin signalling intermediates. We hypothesised that signalling through TBC1D4 is involved in this effect of exercise as it is a common signalling element for insulin and exercise.
Methods: Insulin-regulated glucose metabolism was evaluated in 12 healthy moderately trained young men 4 h after one-legged exercise at basal and during a euglycaemic-hyperinsulinaemic clamp. Vastus lateralis biopsies were taken before and immediately after the clamp.
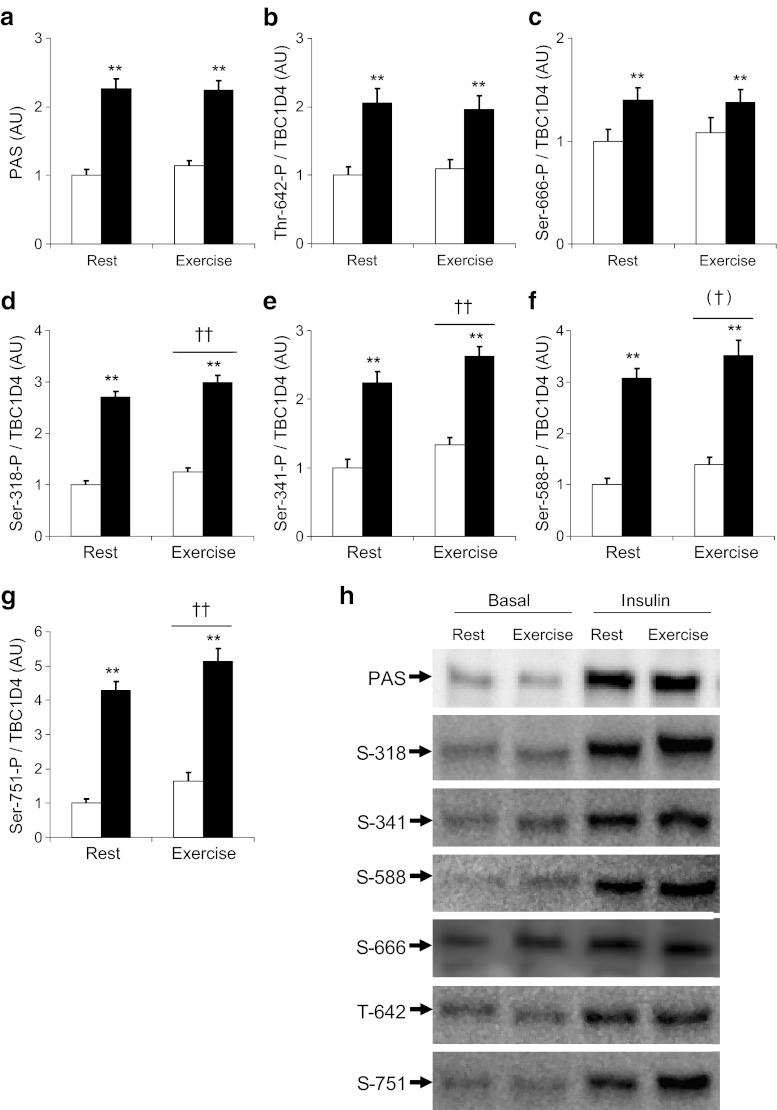
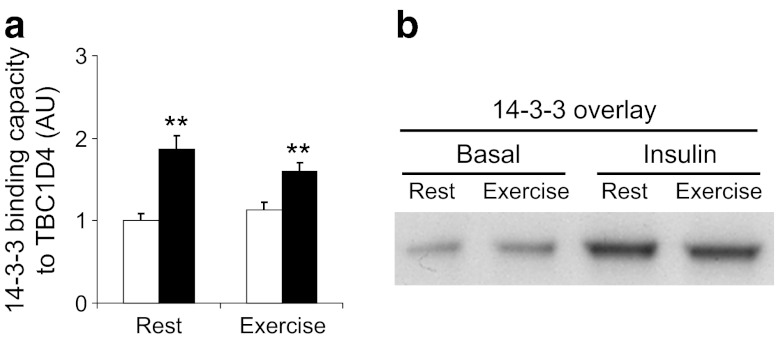
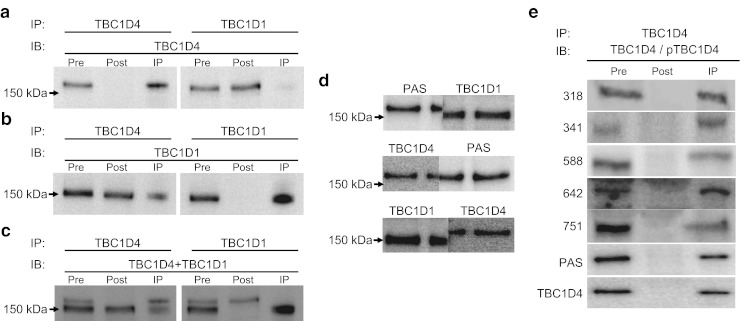
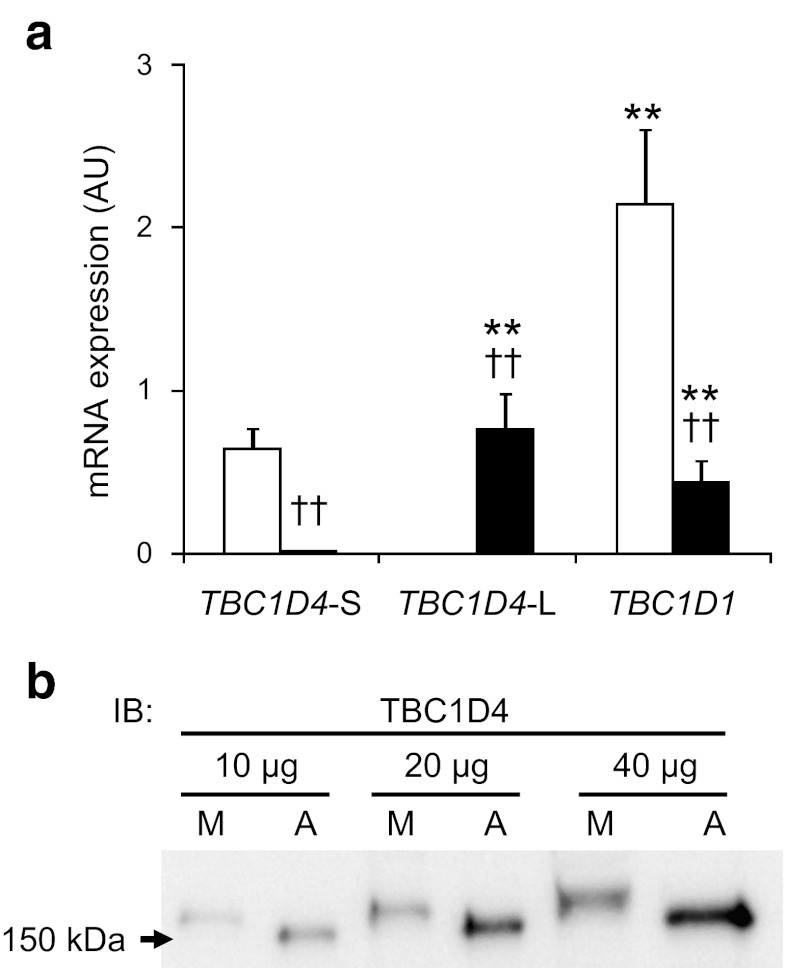
Results: Insulin stimulation increased glucose uptake in both legs, with greater effects (approximately 80%, p < 0.01) in the previously exercised leg. TBC1D4 phosphorylation, assessed using the phospho-AKT (protein kinase B)substrate antibody and phospho- and site-specific antibodies targeting six phosphorylation sites on TBC1D4, increased at similar degrees to insulin stimulation in the previously exercised and rested legs (p < 0.01). However, TBC1D4 phosphorylation on Ser-318, Ser-341, Ser-588 and Ser-751 was higher in the previously exercised leg, both in the absence and in the presence of insulin (p < 0.01; Ser-588, p = 0.09; observed power = 0.39). 14-3-3 binding capacity for TBC1D4 increased equally (p < 0.01) in both legs during insulin stimulation.
Conclusion/interpretation: We provide evidence for site-specific phosphorylation of TBC1D4 in human skeletal muscle in response to physiological hyperinsulinaemia. The data support the idea that TBC1D4 is a nexus for insulin- and exercise-responsive signals that may mediate increased insulin action after exercise.
Figures
References
-
- Garetto LP, Richter EA, Goodman MN, Ruderman NB. Enhanced muscle glucose metabolism after exercise in the rat: the two phases. Am J Physiol Endocrinol Metab. 1984;246:E471–E475. - PubMed
-
- Richter EA, Garetto LP, Goodman MN, Ruderman NB. Enhanced muscle glucose metabolism after exercise: modulation by local factors. Am J Physiol Endocrinol Metab. 1984;246:E476–E482. - PubMed
-
- Mikines KJ, Sonne B, Farrell PA, Tronier B, Galbo H. Effect of physical exercise on sensitivity and responsiveness to insulin in humans. Am J Physiol Endocrinol Metab. 1988;254:E248–E259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
