

Chromatin stability at low concentration depends on histone octamer saturation levels
- PMID: 19254554
- PMCID: PMC2717262
- DOI: 10.1016/j.bpj.2008.10.070
Chromatin stability at low concentration depends on histone octamer saturation levels
Abstract
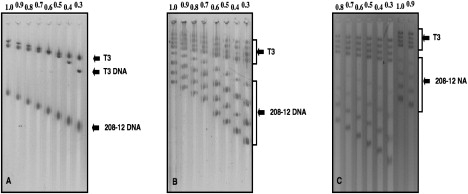
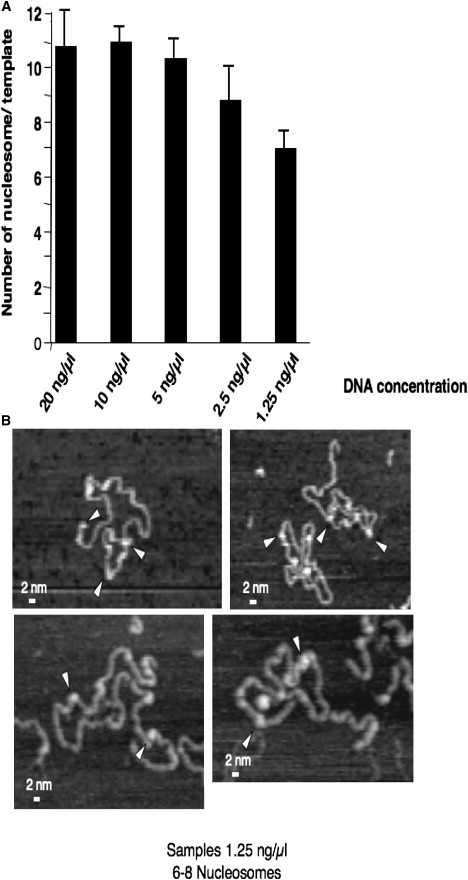
Studies on the stability of nucleosome core particles as a function of concentration have indicated a lower limit of approximately 5 ng/microL, below which the complexes start to spontaneously destabilize. Until recently little information was available on the effect of low concentration on chromatin. Using the well-characterized array of tandemly repeated 5S rDNA reconstituted into chromatin, we have investigated the effect of dilution. In this study, we demonstrate that the stability of saturated nucleosomal arrays and that of nucleosome core particles are within the same order of magnitude, and no significant loss of histones is monitored down to a concentration of 2.5 ng/microL. We observed that levels of subsaturation of the nucleosomal arrays were directly correlated with an increased sensitivity to histone loss, suggesting a shielding effect. The loss of histones from our linear nucleosomal arrays was shown not to be random, with a significant likelihood to occur at the end of the template than toward the center. This observation indicates that centrally located nucleosomes are more stable than those close to the end of the DNA templates. Itis important to take this information into account for the proper design of experiments pertaining to histone composition and the folding ability of chromatin samples.
Figures




References
-
- Schneider R., Grosschedl R. Dynamics and interplay of nuclear architecture, genome organization, and gene expression. Genes Dev. 2007;21:3027–3043. - PubMed
-
- Georgel P.T. Chromatin structure of eukaryotic promoters: a changing perspective. Biochem. Cell Biol. 2002;80:295–300. - PubMed
-
- Hansen J.C. Conformational dynamics of the chromatin fiber in solution: determinants, mechanisms, and functions. Annu. Rev. Biophys. Biomol, Struct. 2002;31:361–392. - PubMed
-
- Woodcock C.L., Dimitrov S. Higher-order structure of chromatin and chromosomes. Curr. Opin. Genet. Dev. 2001;11:130–135. - PubMed
-
- Choudhary P., Varga-Weisz P. ATP-dependent chromatin remodelling: action and reaction. Subcell. Biochem. 2007;41:29–43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
