

Crystal structure of acivicin-inhibited gamma-glutamyltranspeptidase reveals critical roles for its C-terminus in autoprocessing and catalysis
- PMID: 19256527
- PMCID: PMC2668148
- DOI: 10.1021/bi8014955
Crystal structure of acivicin-inhibited gamma-glutamyltranspeptidase reveals critical roles for its C-terminus in autoprocessing and catalysis
Abstract
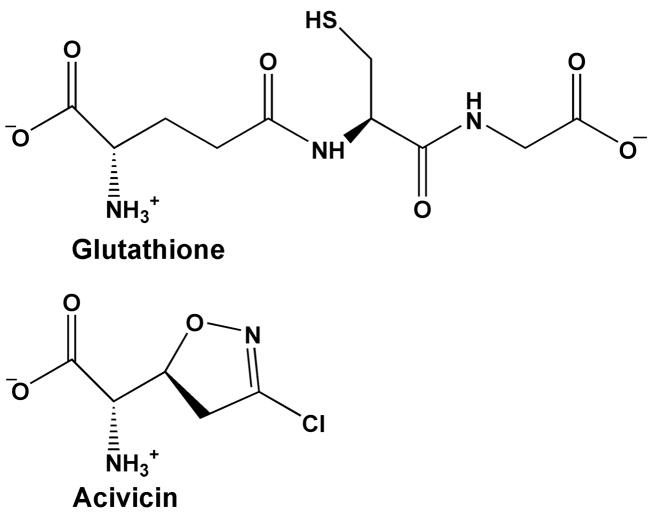
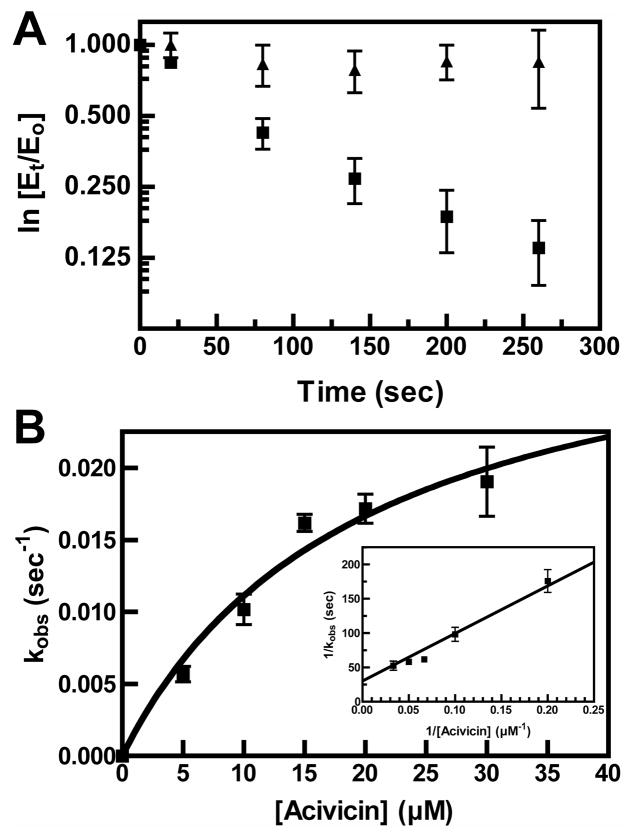
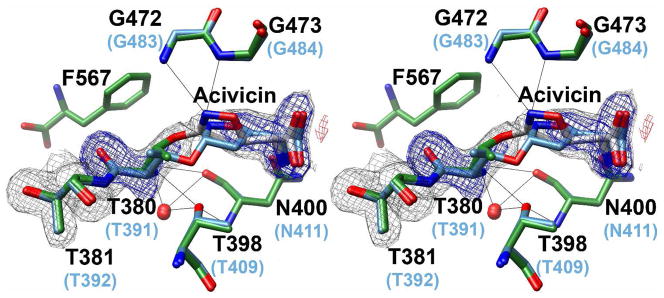
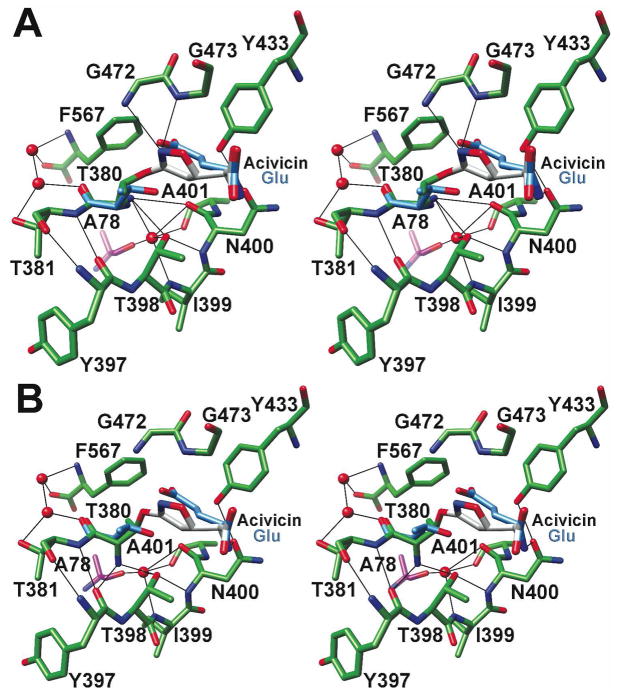
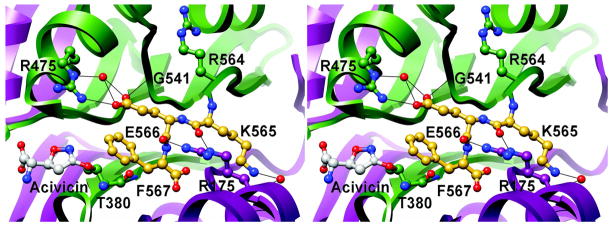
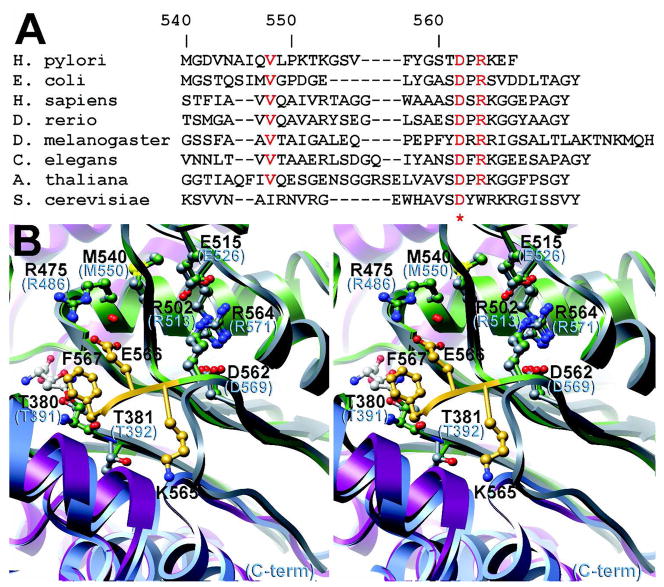
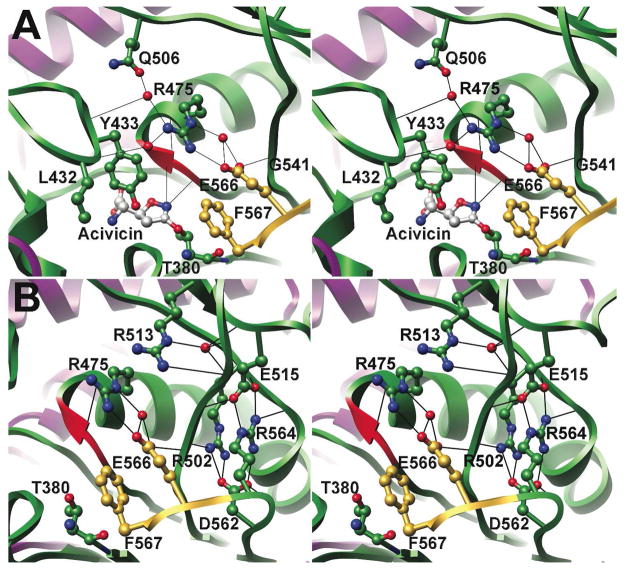
Helicobacter pylori gamma-glutamyltranspeptidase (HpGT) is a general gamma-glutamyl hydrolase and a demonstrated virulence factor. The enzyme confers a growth advantage to the bacterium, providing essential amino acid precursors by initiating the degradation of extracellular glutathione and glutamine. HpGT is a member of the N-terminal nucleophile (Ntn) hydrolase superfamily and undergoes autoprocessing to generate the active form of the enzyme. Acivicin is a widely used gamma-glutamyltranspeptidase inhibitor that covalently modifies the enzyme, but its precise mechanism of action remains unclear. The time-dependent inactivation of HpGT exhibits a hyperbolic dependence on acivicin concentration with k(max) = 0.033 +/- 0.006 s(-1) and K(I) = 19.7 +/- 7.2 microM. Structure determination of acivicin-modified HpGT (1.7 A; R(factor) = 17.9%; R(free) = 20.8%) demonstrates that acivicin is accommodated within the gamma-glutamyl binding pocket of the enzyme. The hydroxyl group of Thr 380, the catalytic nucleophile in the autoprocessing and enzymatic reactions, displaces chloride from the acivicin ring to form the covalently linked complex. Within the acivicin-modified HpGT structure, the C-terminus of the protein becomes ordered with Phe 567 positioned over the active site. Substitution or deletion of Phe 567 leads to a >10-fold reduction in enzymatic activity, underscoring its importance in catalysis. The mobile C-terminus is positioned by several electrostatic interactions within the C-terminal region, most notably a salt bridge between Arg 475 and Glu 566. Mutational analysis reveals that Arg 475 is critical for the proper placement of the C-terminal region, the Tyr 433 containing loop, and the proposed oxyanion hole.
Figures
References
-
- Boanca G, Sand A, Barycki JJ. Uncoupling the enzymatic and autoprocessing activities of Helicobacter pylori gamma-glutamyltranspeptidase. J Biol Chem. 2006;281:19029–19037. - PubMed
-
- Chevalier C, Thiberge JM, Ferrero RL, Labigne A. Essential role of Helicobacter pylori gamma-glutamyltranspeptidase for the colonization of the gastric mucosa of mice. Mol Microbiol. 1999;31:1359–1372. - PubMed
-
- Brannigan JA, Dodson G, Duggleby HJ, Moody PC, Smith JL, Tomchick DR, Murzin AG. A protein catalytic framework with an N-terminal nucleophile is capable of self-activation. Nature. 1995;378:416–419. - PubMed
-
- Suzuki H, Kumagai H. Autocatalytic processing of gamma-glutamyltranspeptidase. J Biol Chem. 2002;277:43536–43543. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
