

Distribution and localization of 5-HT(1A) receptors in the rat lumbar spinal cord after transection and deafferentation
- PMID: 19260781
- PMCID: PMC2828940
- DOI: 10.1089/neu.2008.0640
Distribution and localization of 5-HT(1A) receptors in the rat lumbar spinal cord after transection and deafferentation
Abstract
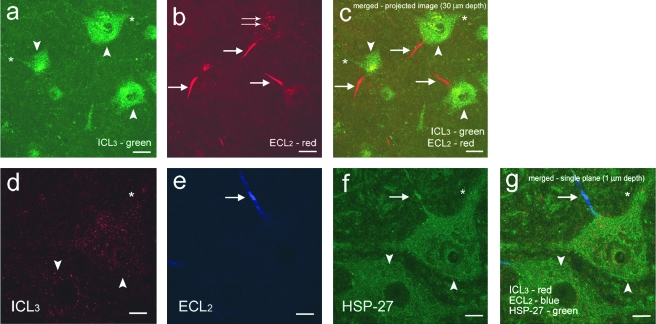
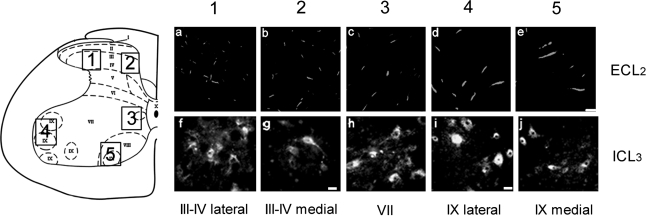
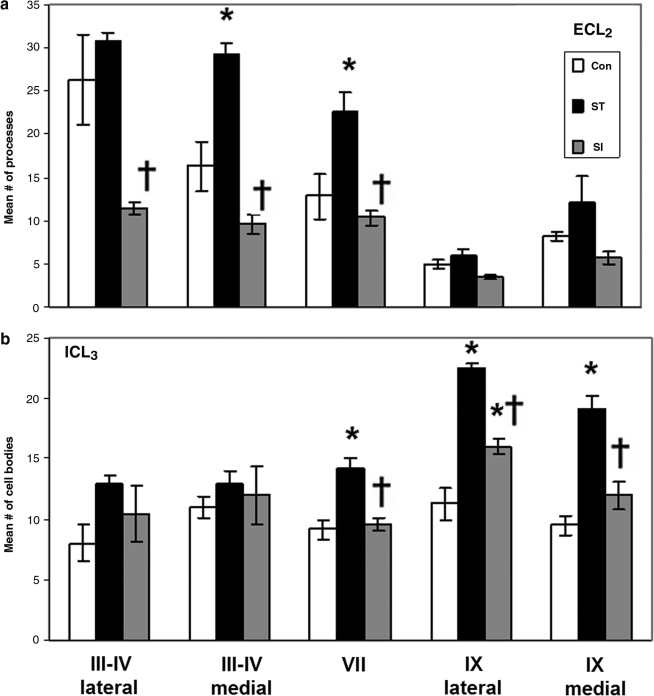
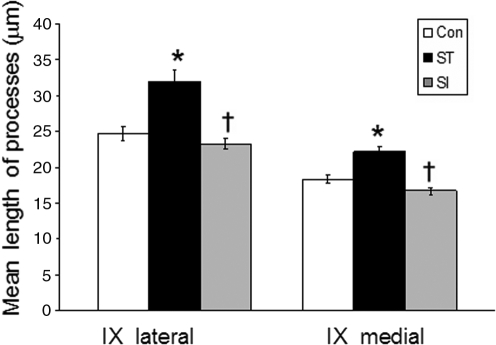
The serotonergic system is highly plastic, capable of adapting to changing afferent information in diverse mammalian systems. We hypothesized that removing supraspinal and/or peripheral input would play an important role in defining the distribution of one of the most prevalent serotonergic receptors, the 5-HT(1A) receptor (R), in the spinal cord. We investigated the distribution of this receptor in response to a complete thoracic (T7-T8) spinal cord transection (eliminating supraspinal input), or to spinal cord isolation (eliminating both supraspinal and peripheral input) in adult rats. Using two antibodies raised against either the second extracellular region (ECL(2)) or the third intracellular region (ICL(3)) of the 5-HT(1A)R, we compared the 5-HT(1A)R levels and distributions in specific laminae of the L3-L5 segments among the control, spinal cord-transected, and spinal cord-isolated groups. Each antibody labeled different populations of 5-HT(1A)R: ECL(2) labeled receptors in the axon hillock, whereas ICL(3) labeled receptors predominantly throughout the soma and proximal dendrites. Spinal cord transection increased the number of ECL(2)-positive cells in the medial region of laminae III-IV and lamina VII, and the mean length of the labeled axon hillocks in lamina IX. The number of ICL(3)-labeled cells was higher in lamina VII and in both the medial and lateral regions of lamina IX in the spinal cord-transected compared to the control group. In contrast, the length and number of ECL(2)-immunolabeled processes and ICL(3)-immunolabeled cells were similar in the spinal cord-isolated and control groups. Combined, these data demonstrate that the upregulation in 5-HT(1A)R that occurs with spinal cord transection alone is dependent on the presence of sensory input.
Figures
Similar articles
-
Serotonin receptors 5-HT1A and 5-HT3 reduce hyperexcitability of dorsal horn neurons after chronic spinal cord hemisection injury in rat.Exp Brain Res. 2003 Mar;149(2):174-86. doi: 10.1007/s00221-002-1352-x. Epub 2003 Jan 25. Exp Brain Res. 2003. PMID: 12610685
-
Locomotor-activated neurons of the cat. I. Serotonergic innervation and co-localization of 5-HT7, 5-HT2A, and 5-HT1A receptors in the thoraco-lumbar spinal cord.J Neurophysiol. 2009 Sep;102(3):1560-76. doi: 10.1152/jn.91179.2008. Epub 2009 Jul 1. J Neurophysiol. 2009. PMID: 19571190 Free PMC article.
-
Induction of c-fos gene expression by spinal cord transection in the rat.Brain Res. 1997 Jul 18;763(1):21-9. doi: 10.1016/s0006-8993(97)00356-9. Brain Res. 1997. PMID: 9272824
-
Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury.Prog Brain Res. 2002;137:257-73. doi: 10.1016/s0079-6123(02)37020-1. Prog Brain Res. 2002. PMID: 12440372 Review.
-
Endogenous neurotrophins and plasticity following spinal deafferentation.Exp Neurol. 2012 May;235(1):70-7. doi: 10.1016/j.expneurol.2010.12.021. Epub 2010 Dec 30. Exp Neurol. 2012. PMID: 21195072 Review.
Cited by
-
Serotonergic pharmacotherapy promotes cortical reorganization after spinal cord injury.Exp Neurol. 2013 Mar;241:84-94. doi: 10.1016/j.expneurol.2012.12.004. Epub 2012 Dec 19. Exp Neurol. 2013. PMID: 23262119 Free PMC article.
-
Recovery of control of posture and locomotion after a spinal cord injury: solutions staring us in the face.Prog Brain Res. 2009;175:393-418. doi: 10.1016/S0079-6123(09)17526-X. Prog Brain Res. 2009. PMID: 19660669 Free PMC article. Review.
-
Activation of 5-HT1A Receptors Normalizes the Overexpression of Presynaptic 5-HT1A Receptors and Alleviates Diabetic Neuropathic Pain.Int J Mol Sci. 2023 Sep 20;24(18):14334. doi: 10.3390/ijms241814334. Int J Mol Sci. 2023. PMID: 37762636 Free PMC article.
-
Changes in functional properties and 5-HT modulation above and below a spinal transection in lamprey.Front Neural Circuits. 2015 Jan 20;8:148. doi: 10.3389/fncir.2014.00148. eCollection 2014. Front Neural Circuits. 2015. PMID: 25653594 Free PMC article.
-
Pronociceptive effect of 5-HT(1A) receptor agonist on visceral pain involves spinal N-methyl-D-aspartate (NMDA) receptor.Neuroscience. 2012 Sep 6;219:243-54. doi: 10.1016/j.neuroscience.2012.05.030. Epub 2012 May 22. Neuroscience. 2012. PMID: 22626644 Free PMC article.
References
-
- Alaimo M. Smith J.L. Roy R.R. Edgerton V.R. EMG activity of slow and fast ankle extensors following spinal cord transection. J. Appl. Physiol. 1984;56:1608–1613. - PubMed
-
- Anden N.E. Haeggendal J. Magnusson T. Rosengren E. The time course of the disappearance of noradrenaline and 5-hydroxytryptamine in the spinal cord after transection. Acta Physiol. Scand. 1964;62:115–118. - PubMed
-
- Andrade R. Malenka R.C. Nicoll R.A. A G protein couples serotonin and GABAB receptors to the same channels in hippocampus. Science. 1986;234:1261–1265. - PubMed
-
- Anthony T.E. Azmitia E.C. Molecular characterization of antipeptide antibodies against the 5-HT1A receptor: evidence for state-dependent antibody binding. Brain Res. Mol. Brain Res. 1997;50:277–284. - PubMed
-
- Antri M. Barthe J.Y. Mouffle C. Orsal D. Long-lasting recovery of locomotor function in chronic spinal rat following chronic combined pharmacological stimulation of serotonergic receptors with 8-OH-DPAT and quipazine. Neurosci. Lett. 2005;384:162–167. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
