

An open-pore structure of the mechanosensitive channel MscL derived by determining transmembrane domain interactions upon gating
- PMID: 19261722
- PMCID: PMC2704598
- DOI: 10.1096/fj.09-129296
An open-pore structure of the mechanosensitive channel MscL derived by determining transmembrane domain interactions upon gating
Abstract
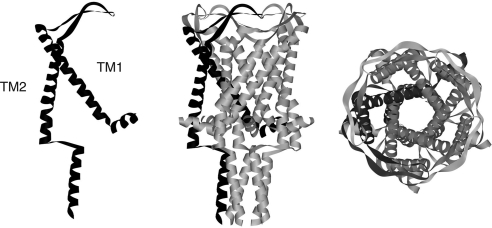
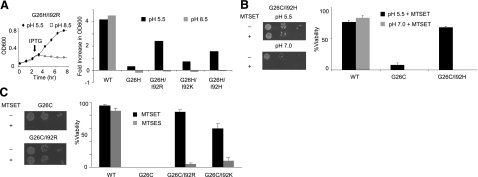
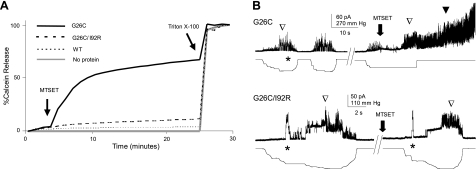
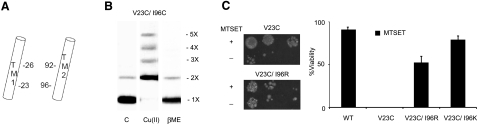
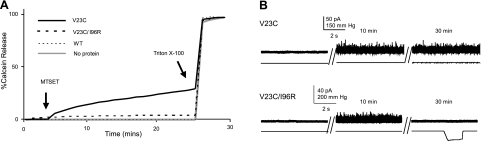
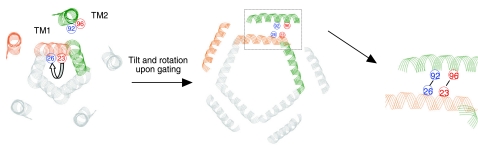
Mechanosensation, the ability to detect mechanical forces, underlies the senses of hearing, balance, touch, and pain, as well as renal and cardiovascular regulation. Although the sensors are thought to be channels, relatively little is known about eukaryotic mechanosensitive channels or their molecular mechanisms. Thus, because of its tractable nature, a bacterial mechanosensitive channel that serves as an in vivo osmotic "emergency release valve," MscL, has become a paradigm of how a mechanosensitive channel can sense and respond to membrane tension. Here, we have determined the structural rearrangements and interactions between transmembrane domains of MscL that occur upon gating. We utilize an electrostatic repulsion test: If two residues approach upon gating we predicted that substituting like-charges at those sites would inhibit gating. The in vivo growth and viability and in vitro vesicular flux and electrophysiological data all support the hypothesis that residues G26 and I92 directly interact upon gating. The resulting model predicted other interacting residues. One of these sets, V23 and I96, was confirmed to truly interact upon gating by disulfide trapping as well as the electrostatic repulsion test. Together, the data strongly suggest a model for structural transitions and residue-residue proximities that occur upon MscL gating.
Figures
Similar articles
-
Electromechanical coupling model of gating the large mechanosensitive ion channel (MscL) of Escherichia coli by mechanical force.Biophys J. 1998 Jun;74(6):2889-902. doi: 10.1016/S0006-3495(98)77995-0. Biophys J. 1998. PMID: 9635742 Free PMC article.
-
An in vivo assay identifies changes in residue accessibility on mechanosensitive channel gating.Proc Natl Acad Sci U S A. 2004 Jul 6;101(27):10161-5. doi: 10.1073/pnas.0402040101. Epub 2004 Jun 28. Proc Natl Acad Sci U S A. 2004. PMID: 15226501 Free PMC article.
-
Electrostatics at the membrane define MscL channel mechanosensitivity and kinetics.FASEB J. 2014 Dec;28(12):5234-41. doi: 10.1096/fj.14-259309. Epub 2014 Sep 15. FASEB J. 2014. PMID: 25223610 Free PMC article.
-
MscL: channeling membrane tension.Pflugers Arch. 2015 Jan;467(1):15-25. doi: 10.1007/s00424-014-1535-x. Epub 2014 May 27. Pflugers Arch. 2015. PMID: 24859800 Free PMC article. Review.
-
Gating the bacterial mechanosensitive channels: MscS a new paradigm?Curr Opin Microbiol. 2004 Apr;7(2):163-7. doi: 10.1016/j.mib.2004.02.006. Curr Opin Microbiol. 2004. PMID: 15063854 Review.
Cited by
-
Improving the Design of a MscL-Based Triggered Nanovalve.Biosensors (Basel). 2013;3(1):171-84. doi: 10.3390/bios3010171. Biosensors (Basel). 2013. PMID: 23678232 Free PMC article.
-
Mechanosensitive channel MscL gating transitions coupling with constriction point shift.Protein Sci. 2024 Apr;33(4):e4965. doi: 10.1002/pro.4965. Protein Sci. 2024. PMID: 38501596 Free PMC article.
-
S. aureus MscL is a pentamer in vivo but of variable stoichiometries in vitro: implications for detergent-solubilized membrane proteins.PLoS Biol. 2010 Dec 7;8(12):e1000555. doi: 10.1371/journal.pbio.1000555. PLoS Biol. 2010. PMID: 21151884 Free PMC article.
-
Mechanical coupling of the multiple structural elements of the large-conductance mechanosensitive channel during expansion.Proc Natl Acad Sci U S A. 2015 Aug 25;112(34):10726-31. doi: 10.1073/pnas.1503202112. Epub 2015 Aug 10. Proc Natl Acad Sci U S A. 2015. PMID: 26261325 Free PMC article.
-
Sensing and responding to membrane tension: the bacterial MscL channel as a model system.Biophys J. 2012 Jul 18;103(2):169-74. doi: 10.1016/j.bpj.2012.06.021. Epub 2012 Jul 17. Biophys J. 2012. PMID: 22853893 Free PMC article. Review.
References
-
- Blount P, Li Y, Moe P C, Iscla I. Mechanosensitive channels gated by membrane tension: bacteria and beyond. Kamkin A, Kiseleva I, editors. New York, NY, USA: Springer Press; Mechanosensitive Ion Channels, Mechanosensitivity in Cells and Tissues, Moscow Academia series. 2008:71–101.
-
- Blount P, Iscla I, Moe P C, Li Y. MscL: The bacterial mechanosensitive channel of large conductance. Hamill O P, editor. St. Louis, MO, USA: Elsevier Press; 2007:202–233.
-
- Moe P, Blount P. Assessment of potential stimuli for mechano-dependent gating of MscL: effects of pressure, tension, and lipid headgroups. Biochemistry. 2005;44:12239–12244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
