

Phosphatidic acid signaling to mTOR: signals for the survival of human cancer cells
- PMID: 19264150
- PMCID: PMC2759177
- DOI: 10.1016/j.bbalip.2009.02.009
Phosphatidic acid signaling to mTOR: signals for the survival of human cancer cells
Abstract
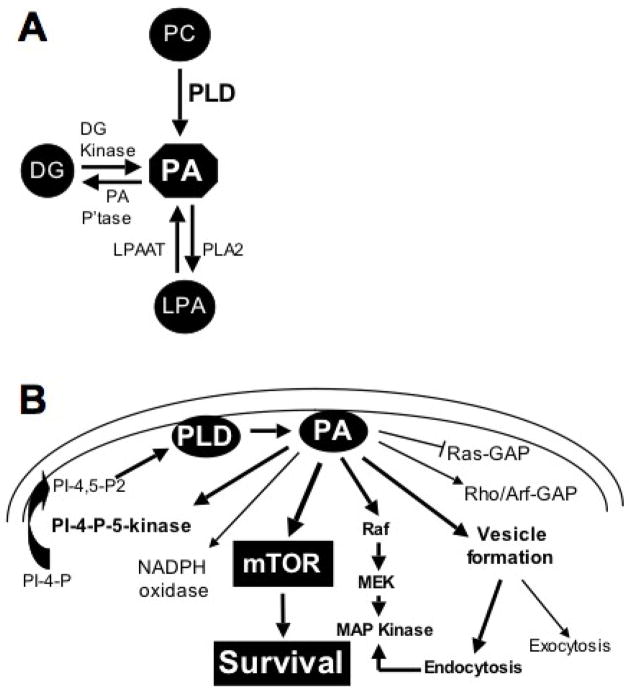
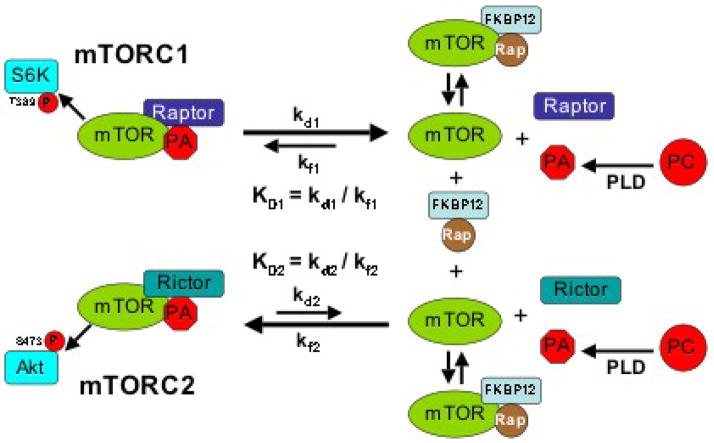
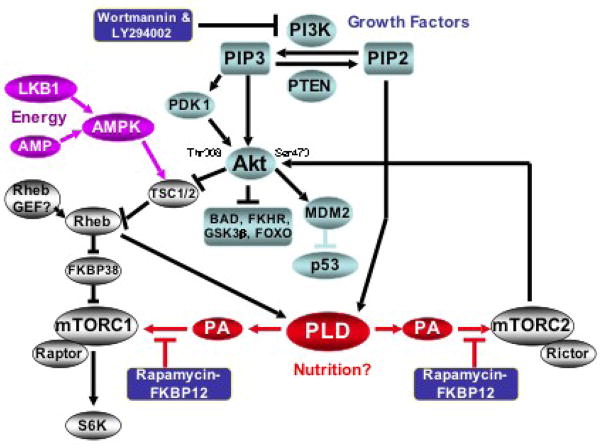
During the past decade elevated phospholipase D (PLD) activity has been reported in virtually all cancers where it has been examined. PLD catalyzes the hydrolysis of phosphatidylcholine to generate the lipid second messenger phosphatidic acid (PA). While many targets of PA signaling have been identified, the most critical target of PA in cancer cells is likely to be mTOR - the mammalian target of rapamycin. mTOR has been widely implicated in signals that suppress apoptotic programs in cancer cells - frequently referred to as survival signals. mTOR exists as two multi-component complexes known as mTORC1 and mTORC2. Recent data has revealed that PA is required for the stability of both mTORC1 and mTORC2 complexes - and therefore also required for the kinase activity of both mTORC1 and mTORC2. PA interacts with mTOR in a manner that is competitive with rapamycin, and as a consequence, elevated PLD activity confers rapamycin resistance - a point that has been largely overlooked in clinical trials involving rapamycin-based strategies. The earliest genetic changes occurring in an emerging tumor are generally ones that suppress default apoptotic programs that likely represent the first line of defense of cancer. Targeting survival signals in human cancers represents a rational anti-cancer therapeutic strategy. Therefore, understanding the signals that regulate PA levels and how PA impacts upon mTOR could be important for developing strategies to de-repress the survival signals that suppress apoptosis. This review summarizes the role of PA in regulating the mTOR-mediated signals that promote cancer cell survival.
Figures
References
-
- Song J, Jiang YW, Foster DA. EGF induces the production of biologically distinguishable diglyceride species from phosphatidylinositol and phosphatidylcholine: evidence for the independent activation of type C and type D phospholipases. Cell Growth Differ. 1994;5:79–85. - PubMed
-
- Motoike T, Bieger S, Wiegandt H, Unsicker K. Induction of phosphatidic acid by fibroblast growth factor in cultured baby hamster kidney fibroblasts. FEBS Lett. 1993;332:164–168. - PubMed
-
- Sa G, Das T. Basic fibroblast growth factor stimulates cytosolic phospholipase A2, phospholipase C-γ1 and phospholipase D through distinguishable signaling mechanisms. Mol Cell Biochem. 1999;198:19–30. - PubMed
-
- Karnam P, Standaert ML, Galloway L, Farese RV. Activation and translocation of Rho and ADP ribosylation factor by insulin in rat adipocytes. Apparent involvement of phosphatidylinositol 3-kinase. J Biol Chem. 1997;272:6136–6140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
