

Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge
- PMID: 19265125
- PMCID: PMC2667713
- DOI: 10.4049/jimmunol.0801814
Tc17, a unique subset of CD8 T cells that can protect against lethal influenza challenge
Abstract
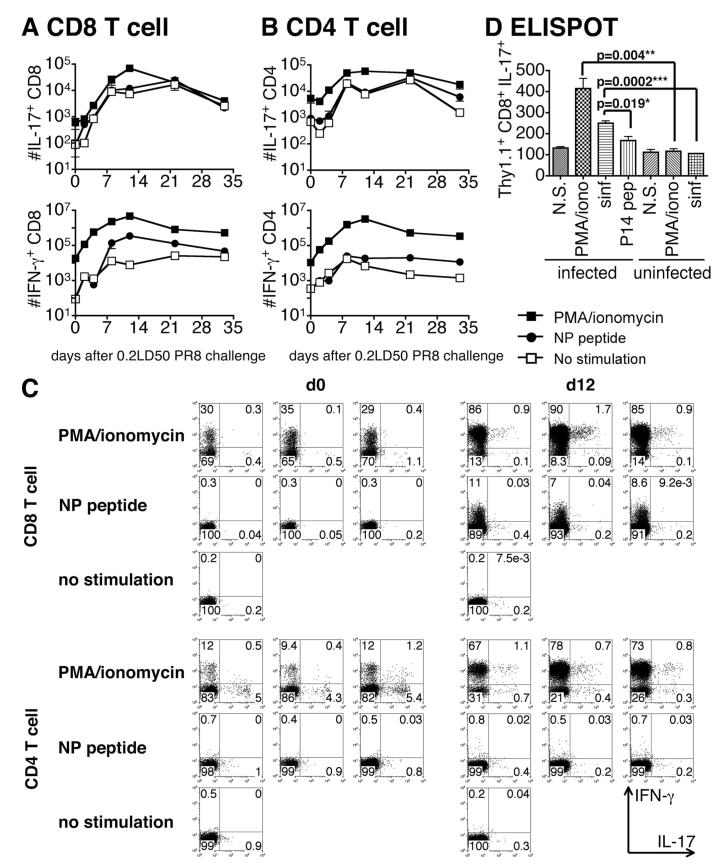
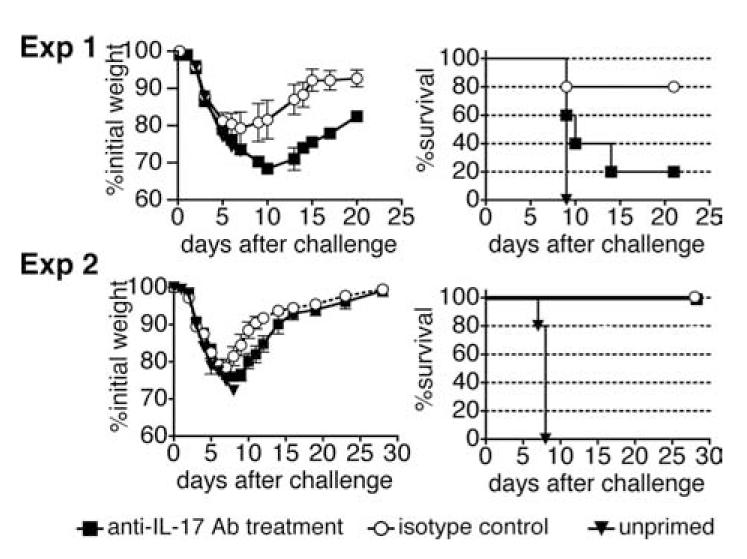
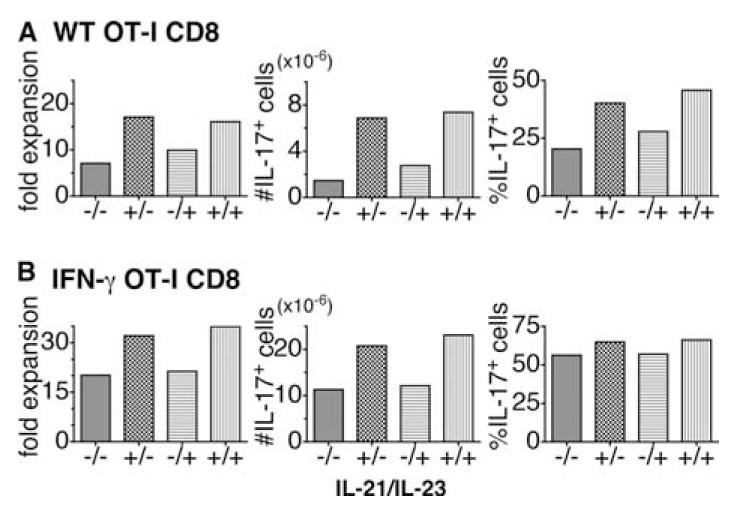
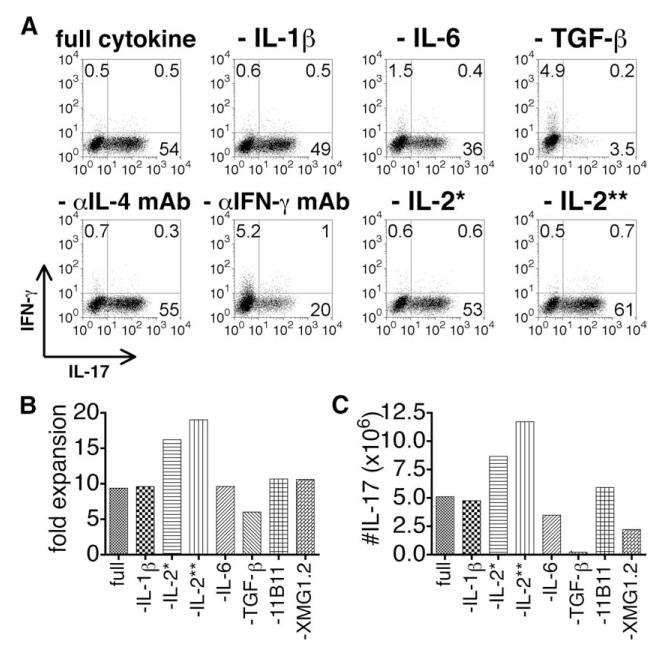
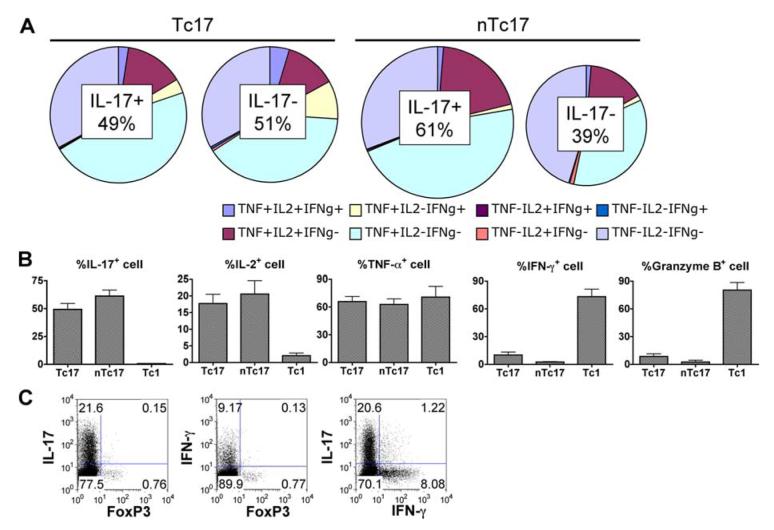
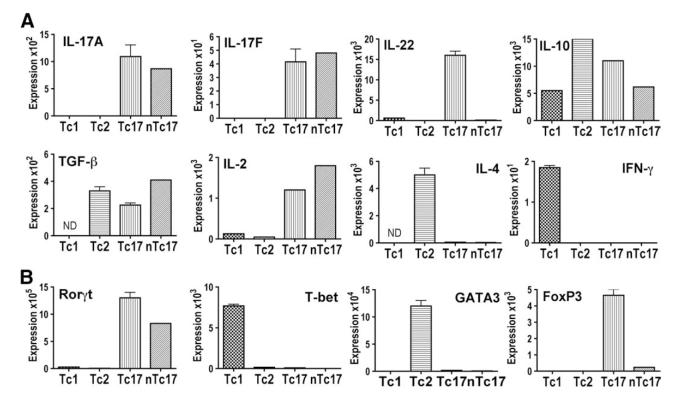
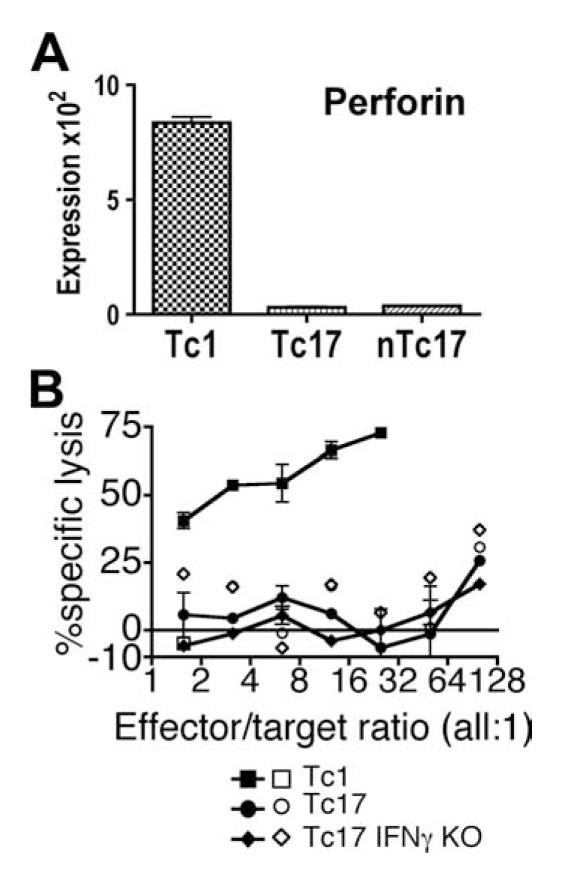
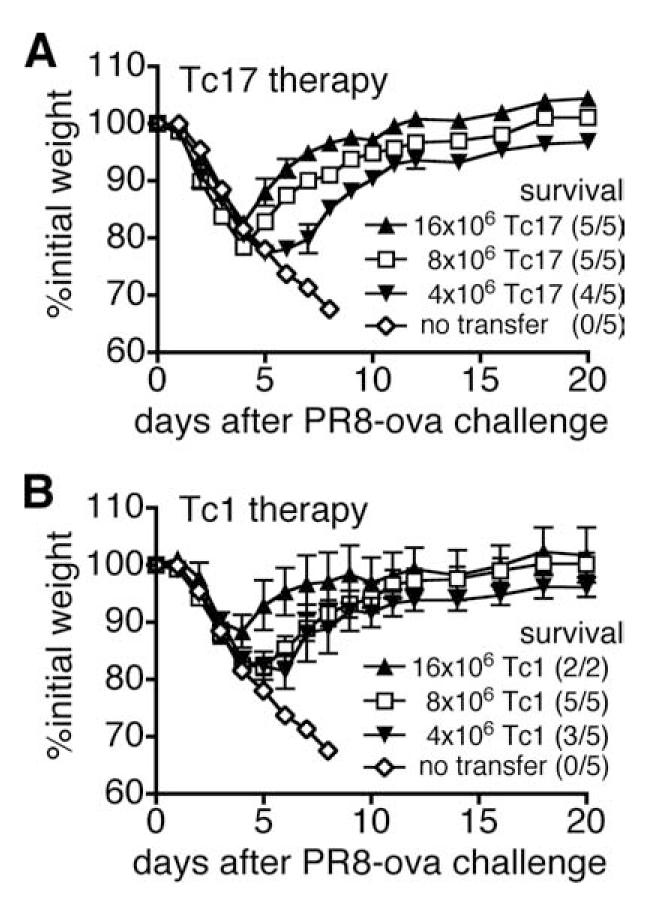
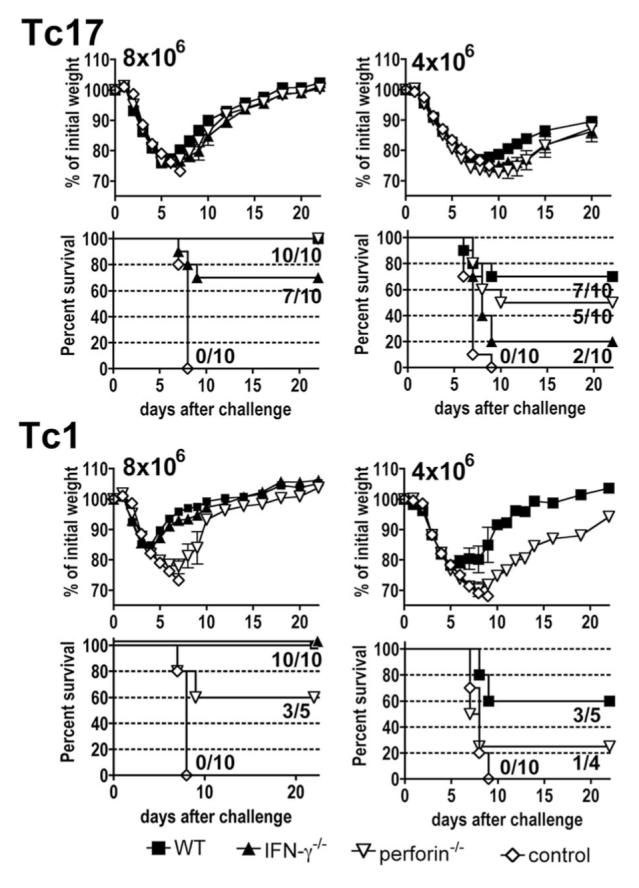
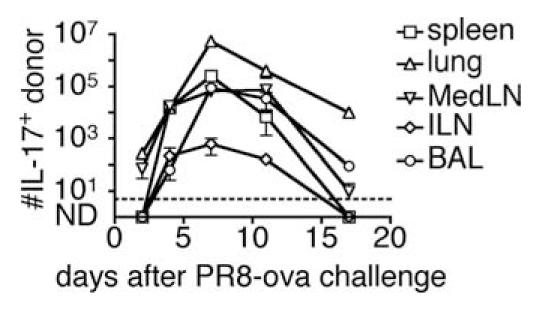
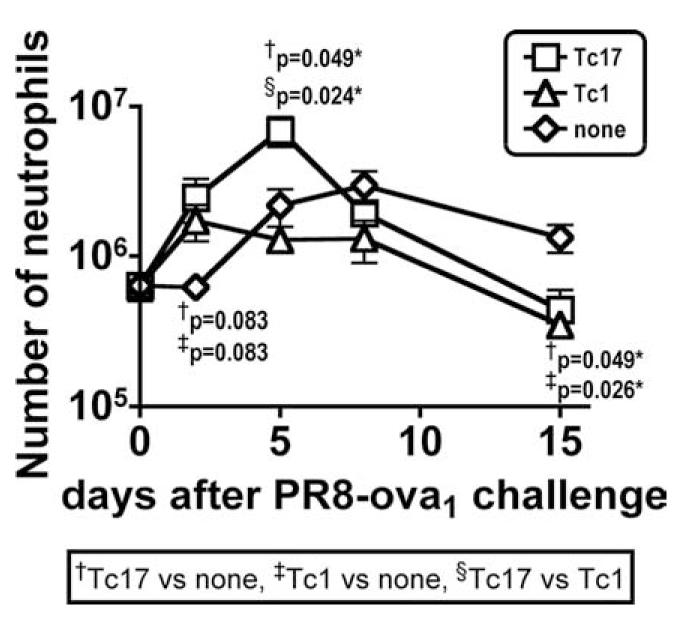
We show here that IL-17-secreting CD4 T (Th)17 and CD8 T (Tc)17 effector cells are found in the lung following primary challenge with influenza A and that blocking Ab to IL-17 increases weight loss and reduces survival. Tc17 effectors can be generated in vitro using naive CD8 T cells from OT-I TCR-transgenic mice. T cell numbers expand 20-fold and a majority secretes IL-17, but little IFN-gamma. Many of the IL-17-secreting cells also secrete TNF and some secrete IL-2. Tc17 are negative for granzyme B, perforin message, and cytolytic activity, in contrast to Tc1 effectors. Tc17 populations express message for orphan nuclear receptor gammat and FoxP3, but are negative for T-bet and GATA-3 transcription factors. The FoxP3-positive, IL-17-secreting and IFN-gamma-secreting cells represent three separate populations. The IFN-gamma-, granzyme B-, FoxP3-positive cells and cells positive for IL-22 come mainly from memory cells and decrease in number when generated from CD44(low) rather than unselected CD8 T cells. Cells of this unique subset of CD8 effector T cells expand greatly after transfer to naive recipients following challenge and can protect them against lethal influenza infection. Tc17 protection is accompanied by greater neutrophil influx into the lung than in Tc1-injected mice, and the protection afforded by Tc17 effectors is less perforin but more IFN-gamma dependent, implying that different mechanisms are involved.
Figures
References
-
- Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989;7:145–173. - PubMed
-
- Swain SL, Bradley LM, Croft M, Tonkonogy S, Atkins G, Weinberg AD, Duncan DD, Hedrick SM, Dutton RW, Huston G. Helper T-cell subsets: phenotype, function and the role of lymphokines in regulating their development. Immunol. Rev. 1991;123:115–144. - PubMed
-
- Seder RA, Paul WE. Acquisition of lymphokine-producing phenotype by CD4+ T cells. Annu. Rev. Immunol. 1994;12:635–673. - PubMed
-
- Seder RA, Boulay JL, Finkelman F, Barbier S, Ben-Sasson SZ, Le Gros G, Paul WE. CD8+ T cells can be primed in vitro to produce IL-4. J. Immunol. 1992;148:1652–1656. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
