

Probing the coupling between proton and electron transfer in photosystem II core complexes containing a 3-fluorotyrosine
- PMID: 19265377
- PMCID: PMC2682732
- DOI: 10.1021/ja808604h
Probing the coupling between proton and electron transfer in photosystem II core complexes containing a 3-fluorotyrosine
Abstract
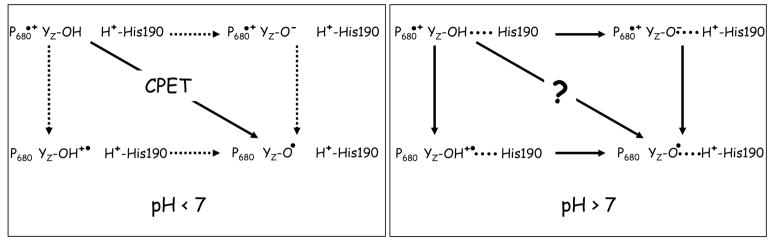
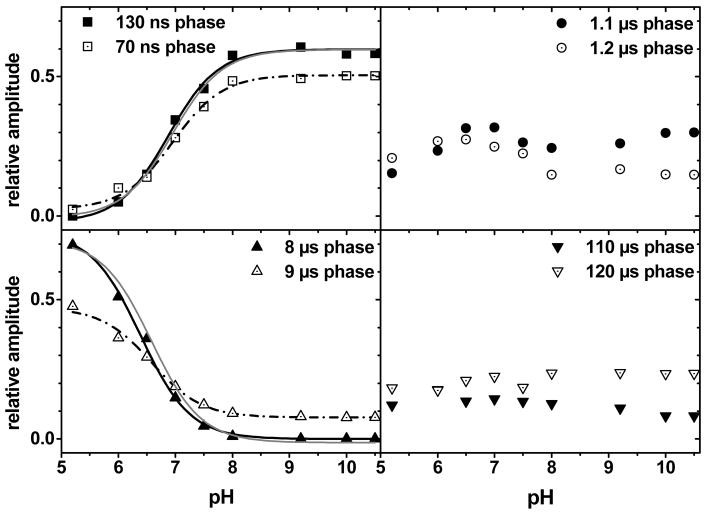
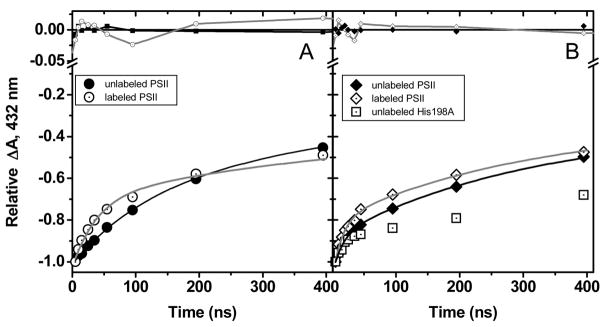
The catalytic cycle of numerous enzymes involves the coupling between proton transfer and electron transfer. Yet, the understanding of this coordinated transfer in biological systems remains limited, likely because its characterization relies on the controlled but experimentally challenging modifications of the free energy changes associated with either the electron or proton transfer. We have performed such a study here in Photosystem II. The driving force for electron transfer from Tyr(Z) to P(680)(*+) has been decreased by approximately 80 meV by mutating the axial ligand of P(680), and that for proton transfer upon oxidation of Tyr(Z) by substituting a 3-fluorotyrosine (3F-Tyr(Z)) for Tyr(Z). In Mn-depleted Photosystem II, the dependence upon pH of the oxidation rates of Tyr(Z) and 3F-Tyr(Z) were found to be similar. However, in the pH range where the phenolic hydroxyl of Tyr(Z) is involved in a H-bond with a proton acceptor, the activation energy of the oxidation of 3F-Tyr(Z) is decreased by 110 meV, a value which correlates with the in vitro finding of a 90 meV stabilization energy to the phenolate form of 3F-Tyr when compared to Tyr (Seyedsayamdost et al. J. Am. Chem. Soc. 2006, 128,1569-1579). Thus, when the phenol of Y(Z) acts as a H-bond donor, its oxidation by P(680)(*+) is controlled by its prior deprotonation. This contrasts with the situation prevailing at lower pH, where the proton acceptor is protonated and therefore unavailable, in which the oxidation-induced proton transfer from the phenolic hydroxyl of Tyr(Z) has been proposed to occur concertedly with the electron transfer to P(680)(*+). This suggests a switch between a concerted proton/electron transfer at pHs < 7.5 to a sequential one at pHs > 7.5 and illustrates the roles of the H-bond and of the likely salt-bridge existing between the phenolate and the nearby proton acceptor in determining the coupling between proton and electron transfer.
Figures
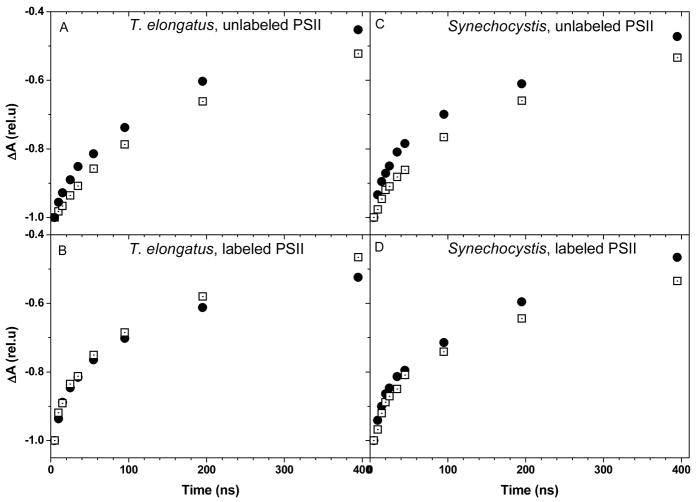
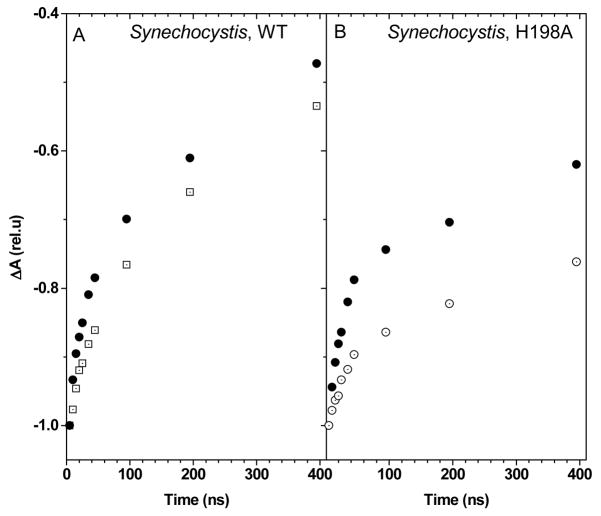
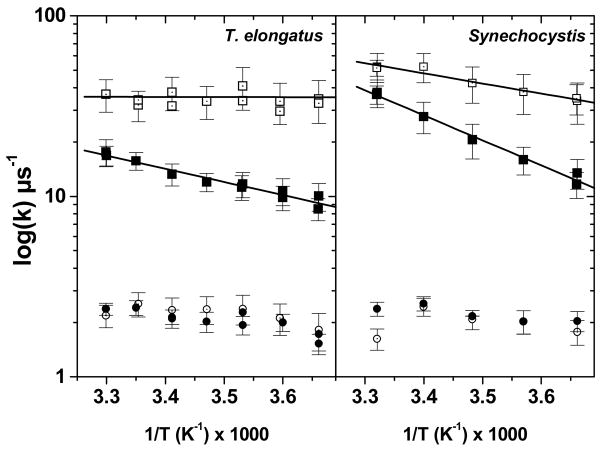
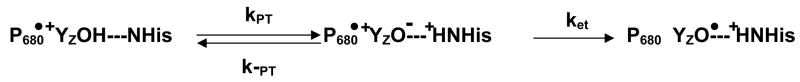
References
-
- Diner BA, Rappaport F. Annu Rev Plant Biol. 2002;53:551–580. - PubMed
-
- Rappaport F, Diner BA. Coord Chem Rev. 2008;252:259–272.
-
- Dau H, Haumann M. Coord Chem Rev. 2008;252:273–295.
-
- Rappaport F, Guergova-Kuras M, Nixon PJ, Diner BA, Lavergne J. Biochemistry. 2002;41:8518–8527. - PubMed
-
- Cuni A, Xiong L, Sayre RT, Rappaport F, Lavergne J. Phys Chem Chem Phys. 2004;6:4825–4831.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
