

Epstein-Barr Nuclear Antigen 1 modulates replication of oriP-plasmids by impeding replication and transcription fork migration through the family of repeats
- PMID: 19265546
- PMCID: PMC2654434
- DOI: 10.1186/1743-422X-6-29
Epstein-Barr Nuclear Antigen 1 modulates replication of oriP-plasmids by impeding replication and transcription fork migration through the family of repeats
Abstract
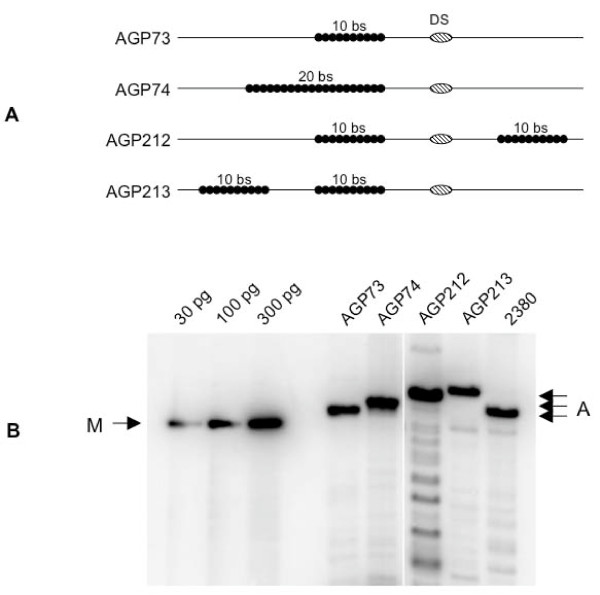
Background: Epstein-Barr virus is replicated once per cell-cycle, and partitioned equally in latently infected cells. Both these processes require a single viral cis-element, termed oriP, and a single viral protein, EBNA1. EBNA1 binds two clusters of binding sites in oriP, termed the dyad symmetry element (DS) and the family of repeats (FR), which function as a replication element and partitioning element respectively. Wild-type FR contains 20 binding sites for EBNA1.
Results: We, and others, have determined previously that decreasing the number of EBNA1-binding sites in FR increases the efficiency with which oriP-plasmids are replicated. Here we demonstrate that the wild-type number of binding sites in FR impedes the migration of replication and transcription forks. Further, splitting FR into two widely separated sets of ten binding sites causes a ten-fold increase in the efficiency with which oriP-plasmids are established in cells expressing EBNA1. We have also determined that EBNA1 bound to FR impairs the migration of transcription forks in a manner dependent on the number of EBNA1-binding sites in FR.
Conclusion: We conclude that EBNA1 bound to FR regulates the replication of oriP-plasmids by impeding the migration of replication forks. Upon binding FR, EBNA1 also blocks the migration of transcription forks. Thus, in addition to regulating oriP replication, EBNA1 bound to FR also decreases the probability of detrimental collisions between two opposing replication forks, or between a transcription fork and a replication fork.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
