

doi: 10.1101/gad.1770009.
Pattern formation via small RNA mobility
Affiliations
- PMID: 19270155
- PMCID: PMC2658522
- DOI: 10.1101/gad.1770009
Item in Clipboard
Pattern formation via small RNA mobility
Genes Dev.
.
Abstract
MicroRNAs and trans-acting siRNAs (ta-siRNAs) have important regulatory roles in development. Unlike other developmentally important regulatory molecules, small RNAs are not known to act as mobile signals during development. Here, we show that low-abundant, conserved ta-siRNAs, termed tasiR-ARFs, move intercellularly from their defined source of biogenesis on the upper (adaxial) side of leaves to the lower (abaxial) side to create a gradient of small RNAs that patterns the abaxial determinant AUXIN RESPONSE FACTOR3. Our observations have important ramifications for the function of small RNAs and suggest they can serve as mobile, instructive signals during development.
Figures
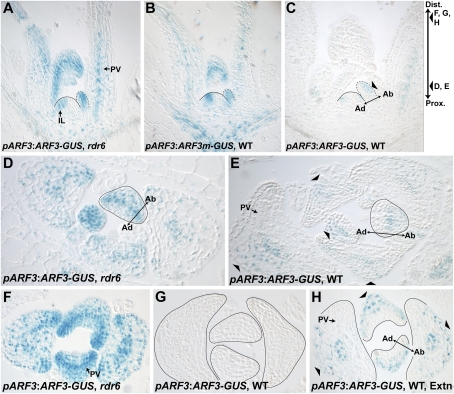
Differential tasiR-ARF activity across the adaxial-abaxial axis of leaf primordia. (A) In rdr6-15, pARF3:ARF3-GUS is expressed throughout leaf primordia and below the SAM. (IL) Incipient leaf; (PV) provasculature. (B) The tasiR-ARF insensitive pARF3:ARF3m-GUS reporter in wild type is expressed similarly. (C) pARF3:ARF3-GUS expression in wild type is reduced and abaxially restricted (arrowhead). (D,E) Transverse sections as indicated in C, showing pARF3:ARF3-GUS expression throughout primordia in rdr6-15 (D), but abaxially restricted in wild type (E). (F,G) Distal sections as indicated in C showing strong pARF3:ARF3-GUS activity throughout rdr6-15 leaves (F), but no activity in wild type (G). (H) Section identical to G but with extended GUS staining time shows abaxial pARF3:ARF3-GUS activity (arrowheads).
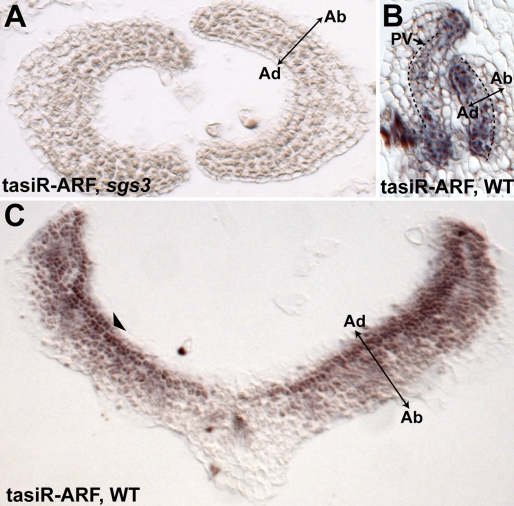
tasiR-ARFs differentially accumulate along the adaxial-abaxial axis. (A) In situ hybridization in an sgs3 background using the tasiR-ARF LNA probe yields no hybridization signal above background, indicating the probe's specificity for tasiR-ARFs. (B) tasiR-ARFs accumulate on the adaxial side of young primordia near the SAM. Note their predominance in the margins and provasculature (PV, arrow). (C) In distal regions of older leaves, tasiR-ARFs accumulate strongest adaxially (arrowhead) and form a gradient that dissipates toward the abaxial side of the leaf.
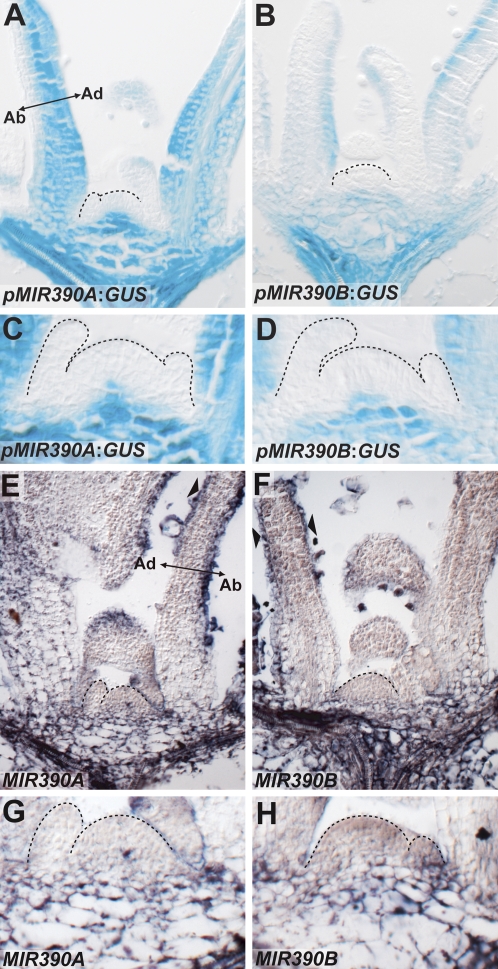
Absence of MIR390 precursor transcripts in the meristem and young leaf primordia. (A,B) Reporters for MIR390A (A) and MIR390B (B) show transcriptional activity for these loci in the vasculature and pith region beneath the SAM, but not within the meristem or youngest leaf primordia. In older leaf primordia, MIR390A:GUS is expressed more broadly than MIR390B:GUS. (C,D) Close-ups illustrating the lack of MIR390A:GUS (C) and MIR390B:GUS (D) reporter activity in the SAM and young leaf primordia. (E,F) In situ hybridizations show accumulation of MIR390A (E) and MIR390B (F) precursor transcripts in patterns similar to the expression patterns of their respective reporters. (G,H) Close-ups of meristem regions showing MIR390A (G) and MIR390B (H) accumulation beneath the SAM but not within the meristem or young leaf primordia. Note, sense probes for MIR390A and MIR390B yield no hybridization signal (Suppelemental Fig. S3).
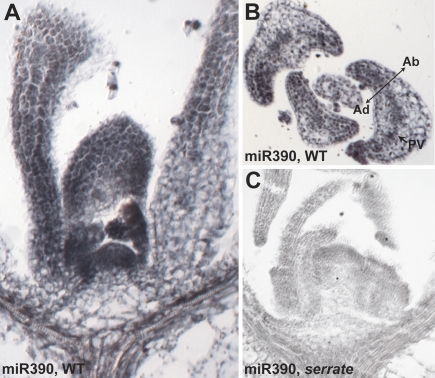
Mature miR390 accumulates throughout the shoot apex. (A) In situ hybridization shows miR390 accumulates robustly in the SAM and young primordia as well as in the vasculature and pith region below the SAM. (B) In leaves, miR390 accumulates throughout the leaf, but especially within the margins and provasculature (PV, arrow). (C) In serrate-2, the miR390 hybridization signal is barely detectable.
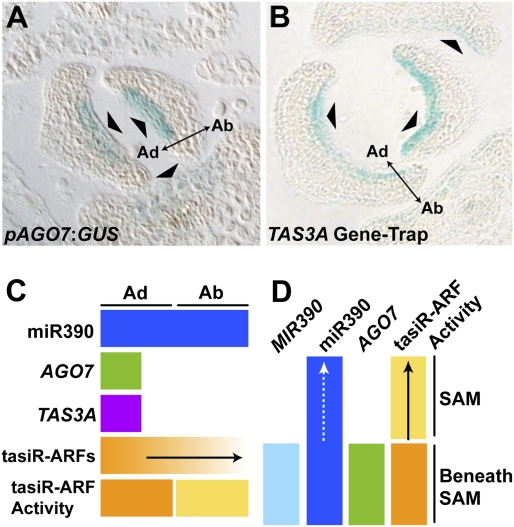
Localized expression of AGO7 and TAS3A restricts the domain of tasiR-ARF biogenesis. (A) AGO7 expression is restricted to a central region of the adaxial-most cell layers of leaves (arrowheads). (B) Expression of TAS3A is similarly adaxially restricted in leaves (arrowheads). (C,D) Summary of the different localization patterns of tasiR-ARF pathway components and a model of small RNA mobility (arrows) in leaves (C) and the SAM (D).
References
-
- Adenot X., Elmayan T., Lauressergues D., Boutet S., Bouche N., Gasciolli V., Vaucheret H. DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol. 2006;16:927–932. - PubMed
-
- Allen E., Xie Z., Gustafson A., Carrington J. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources