

Review
doi: 10.1242/dev.014423.
Molecules and mechanisms of dendrite development in Drosophila
Affiliations
- PMID: 19270170
- PMCID: PMC2685926
- DOI: 10.1242/dev.014423
Item in Clipboard
Review
Molecules and mechanisms of dendrite development in Drosophila
Development.
2009 Apr.
Abstract
Neurons are one of the most morphologically diverse cell types, in large part owing to their intricate dendrite branching patterns. Dendrites are structures that are specialized to receive and process inputs in neurons, thus their specific morphologies reflect neural connectivity and influence information flow through circuits. Recent studies in Drosophila on the molecular basis of dendrite diversity, dendritic guidance, the cell biology of dendritic branch patterning and territory formation have identified numerous intrinsic and extrinsic cues that shape diverse features of dendrites. As we discuss in this review, many of the mechanisms that are being elucidated show conservation in diverse systems.
Figures
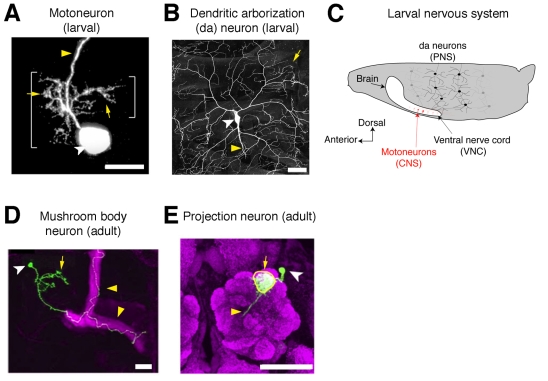
Diverse morphologies of Drosophila dendrites. (A) A
Drosophila RP2 motoneuron projects its dendritic arbor within the
ventral nerve cord of the embryonic CNS. Adapted with permission from Ou et
al. (Ou et al., 2008). Yellow
arrows indicate dendrites, yellow arrowheads indicate axons, and cell bodies
are indicated by white arrowheads in this and subsequent panels. (B)
The dendrites of a highly branched class IV dendritic arborization (da) neuron
of a third-instar Drosophila larva. Image reproduced with permission
from Matthews et al. (Matthews et al.,
2007). (C) Schematic of a Drosophila larva showing
the location of da neurons in the peripheral nervous system (PNS) and
motoneuron cell bodies (red) within the central nervous system (CNS) (not all
segments or cells are shown). Anterior is to the left and dorsal is up.
(D) A mushroom body neuron (green) elaborates dendrites near the cell
body. Image reproduced with permission from Zhu et al.
(Zhu, S. et al., 2006).
(E) A single projection neuron projects its dendrites to a single
glomerulus (outlined in yellow) within the antennal lobe (magenta). Image
reproduced with permission from Komiyama and Luo
(Komiyama and Luo, 2007).
Scale bars: 10 μm in A; 50 μm in B,E; 20 μm in D.
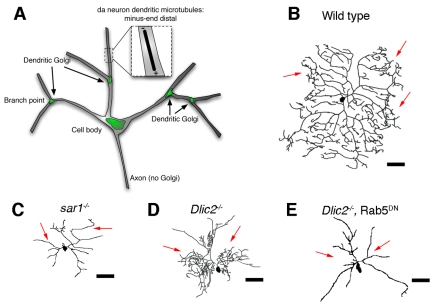
Organelle trafficking and dendrite morphogenesis. (A)
Schematic of Golgi distribution in Drosophila da neurons. Golgi
outposts are localized to dendritic branch points and excluded from the axon.
The predominant minus-end distal arrangement of microtubules in these
dendrites is also shown. (B-E) Dendritic morphologies of wild-type and
mutant class IV Drosophila da neurons. Arrows indicate dendrites. (B)
Tracing of a wild-type class IV da neuron. (C) Tracing of a sar1
mutant class IV da neuron that shows reduced branch complexity. Sar1 is
involved in the formation of COPII vesicles during trafficking from ER to the
Golgi. (D) Tracing of a class IV da neuron with a mutation in dynein light
intermediate chain (Dlic2) showing reduced dendrite length and
redistribution of branches to areas nearer to the cell body (shown in black).
Dlic2 is a component of the dynein complex, a minus-end-directed microtubule
motor. (E) Tracing of a class IV da neuron expressing a dominant-negative Rab5
[Rab5(S43N)]. Dominant-negative Rab5 abrogates the proximal hyperbranching
phenotype of Dlic2 mutations. Rab5 is a GTPase that functions in
early endocytosis. Tracings in B,C adapted with permission from Ye et al.
(Ye et al., 2007). Tracings
in D,E adapted with permission from Satoh et al.
(Satoh et al., 2008). Scale
bars: 75 μm.
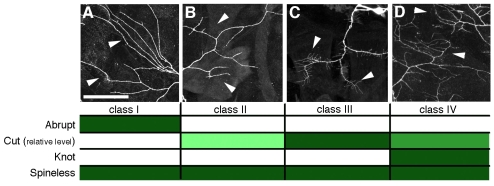
Diversity of da neuron morphology and transcription factor
expression. (A-D) Dendritic arbors of class I, II, III and IV da
neurons (left to right). Arrowheads indicate regions of arbors that exemplify
class-specific branching complexity. Cells are classified according to
increasing arbor complexity. The expression status of transcription factors
Cut, Knot, Abrupt and Spineless is listed below each morphological class.
Filled boxes indicate expression, white boxes indicate no detectable
expression. Progressively higher levels of Cut expression are indicated by
progressively darker shadings (the degree of shading is not intended to
indicate relative levels among the different transcription factors). Images in
A-C reproduced with permission from Matthews et al.
(Matthews et al., 2007). Scale
bar: 50 μm.
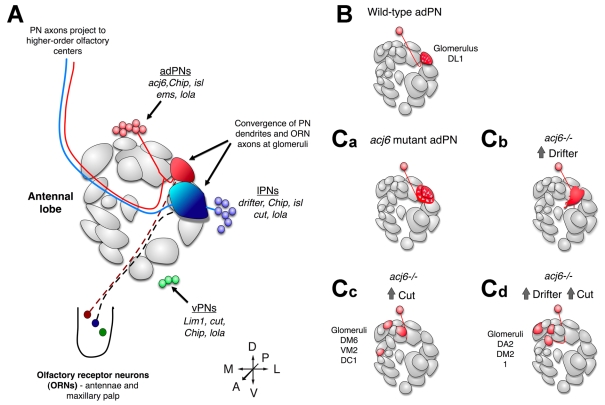
Transcriptional control of dendritic targeting in the
Drosophila antennal lobe. (A) Schematic organization of
the Drosophila antennal lobe (AL). For simplicity, only a subset of
glomeruli are shown. Projection neurons (PNs) from anterodorsal (adPNs, red),
lateral (lPNs, blue) and ventral (vPNs, green) lineages project dendrites to
glomeruli where they connect with olfactory receptor neuron (ORN) axons (a vPN
projection is not shown here). PN axons extend to higher-order olfactory
centers in the brain. Transcription factors discussed in this review are
shown. The schematic of glomerular organization is based on data from Couto et
al. and is adapted with permission (Couto
et al., 2005). (isl is also known as tup -
Flybase.) (B-Cd) Cell-autonomous alterations in transcription factor
expression redirect dendrite targeting. (B) Wild-type DL1 adPN (red) dendrites
normally target to the DL1 glomerulus (shaded in red in the AL). adPNs express
acj6 but not drifter or cut. (Ca-Cd) Dendrite
targeting of genetically manipulated DL1 adPNs. (Ca) acj6 mutants
extend dendrites outside their normal glomerulus. (Cb) acj6 mutant
DL1 adPNs forced to express Drifter partially mistarget to more anterior
glomeruli. (Cc) acj6 mutant DL1 adPNs forced to express Cut target
medial adPN glomeruli. (Cd) Expression of both Drifter and Cut in
acj6 mutant DL1 adPNs results in mistargeting of dendrites to medial
lPN glomeruli. Schematic based on published data
(Komiyama et al., 2003;
Komiyama and Luo, 2007).
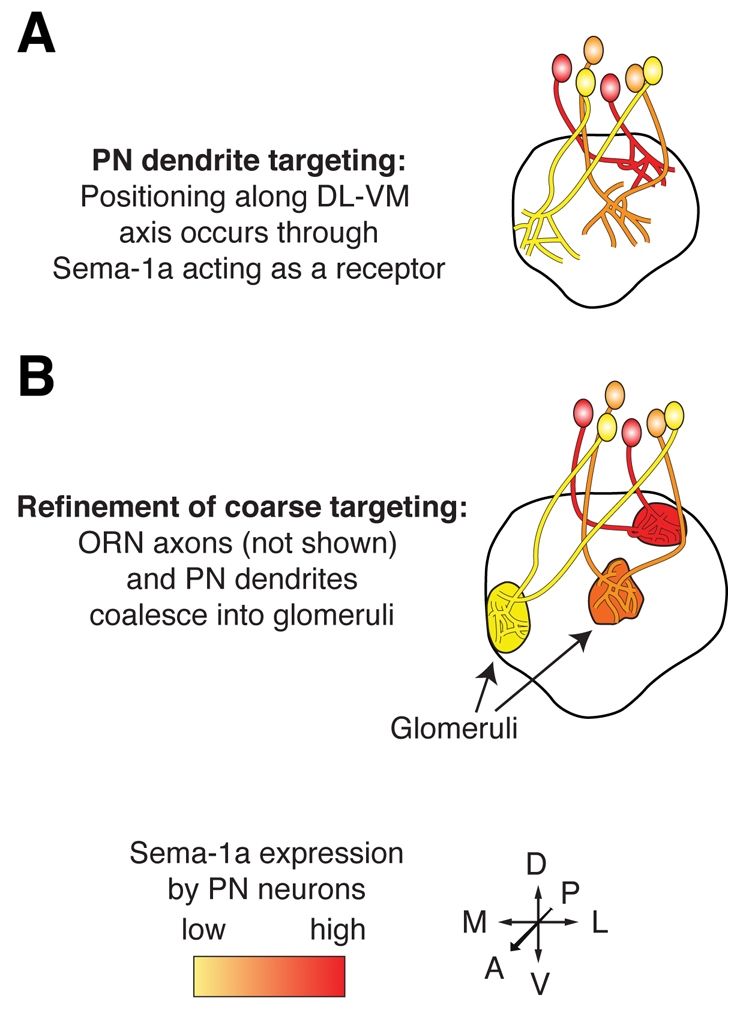
Graded expression of Sema-1a directs projection neuron dendrite
targeting. (A) Projection neurons (PNs) that express the highest
levels of Sema-1a (red) form protoglomeruli at the most dorsolateral (DL)
regions of the Drosophila antennal lobe (AL), whereas those that
express lower levels target to more ventromedial (VM) regions (orange and
yellow). (B) Olfactory receptor neuron (ORN) axons (not shown) and PN
dendrites coalesce into mature glomeruli through axon-dendrite and
dendrite-dendrite interactions (see text). Manipulation of Sema-1a levels in
PNs causes autonomous switches in dendrite targeting [see text and Komiyama et
al. (Komiyama et al., 2007)].
Adapted with permission from Komiyama et al.
(Komiyama et al., 2007).
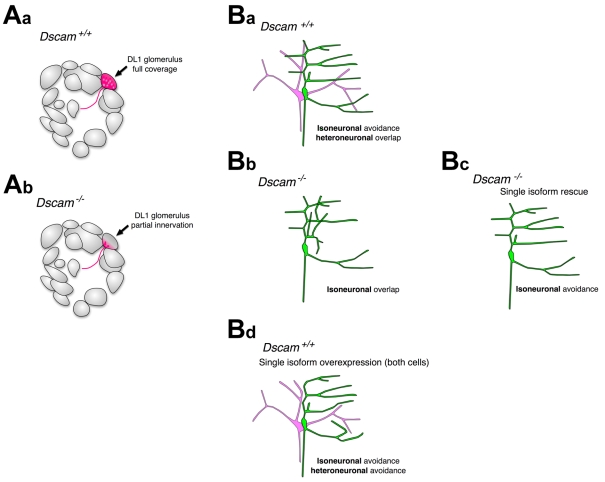
Role of Dscam in dendrite self-avoidance. (Aa)
Schematic of a wild-type projection neuron (PN) targeting the DL1 glomerulus
in the Drosophila antennal lobe. Dendrites project throughout the
entire glomerulus (red). (Ab) In a Dscam mutant DL1 clone,
dendrites fail to completely innervate their target glomerulus (coverage area
in red). Schematic based on published data
(Zhu, H. et al., 2006).
(Ba-Bd) Dscam control of self-avoidance in Drosophila da
neurons. Schematic of two da neurons and their dendritic arbors (green and
pink). (Ba) In a wild-type da neuron, dendrites of each cell self-avoid, but
overlap with non-sister dendrites. Isoneuronal refers to the behavior of
dendrites from the same cell (sister dendrites) and heteroneuronal refers to
the behavior of dendrites from different cells. (Bb-Bd) Schematics of
dendritic phenotypes observed in Dscam mutant animals. (Bb) In a
Dscam mutant, the dendrites of each cell fail to self-avoid and
overlap extensively throughout their arbors. (Bc) Single isoforms expressed in
one Dscam mutant cell (green) rescue self-avoidance. (Bd)
Overexpression of a single isoform in two cells with dendrites that normally
overlap leads to repulsion between branches (heteroneuronal avoidance).
Schematics based on published data (Hughes
et al., 2007; Matthews et al.,
2007; Soba et al.,
2007).
References
-
- Agarwala, K. L., Nakamura, S., Tsutsumi, Y. and Yamakawa, K. (2000). Down syndrome cell adhesion molecule DSCAM mediates homophilic intercellular adhesion. Brain Res. Mol. Brain Res. 79, 118-126. - PubMed
-
- Agarwala, K. L., Ganesh, S., Amano, K., Suzuki, T. and Yamakawa, K. (2001). DSCAM, a highly conserved gene in mammals, expressed in differentiating mouse brain. Biochem. Biophys. Res. Commun. 281, 697-705. - PubMed
-
- Ainsley, J. A., Pettus, J. M., Bosenko, D., Gerstein, C. E., Zinkevich, N., Anderson, M. G., Adams, C. M., Welsh, M. J. and Johnson, W. A. (2003). Enhanced locomotion caused by loss of the Drosophila DEG/ENaC protein Pickpocket1. Curr. Biol. 13, 1557-1563. - PubMed
-
- Aizawa, H., Hu, S. C., Bobb, K., Balakrishnan, K., Ince, G., Gurevich, I., Cowan, M. and Ghosh, A. (2004). Dendrite development regulated by CREST, a calcium-regulated transcriptional activator. Science 303, 197-202. - PubMed
-
- Alvarez, V. A. and Sabatini, B. L. (2007). Anatomical and physiological plasticity of dendritic spines. Annu. Rev. Neurosci. 30, 79-97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
