

Defects in myelination, paranode organization and Purkinje cell innervation in the ether lipid-deficient mouse cerebellum
- PMID: 19270340
- PMCID: PMC2722228
- DOI: 10.1093/hmg/ddp110
Defects in myelination, paranode organization and Purkinje cell innervation in the ether lipid-deficient mouse cerebellum
Abstract
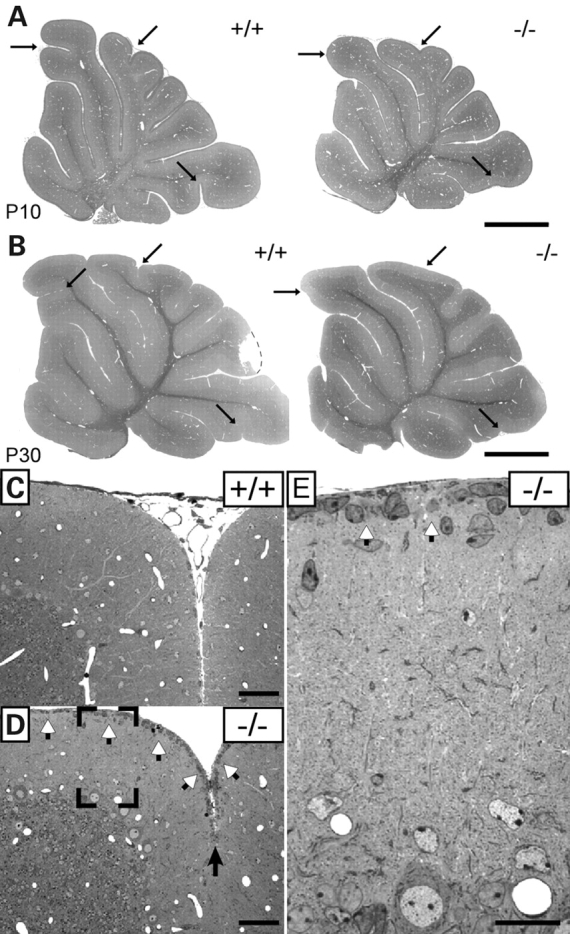
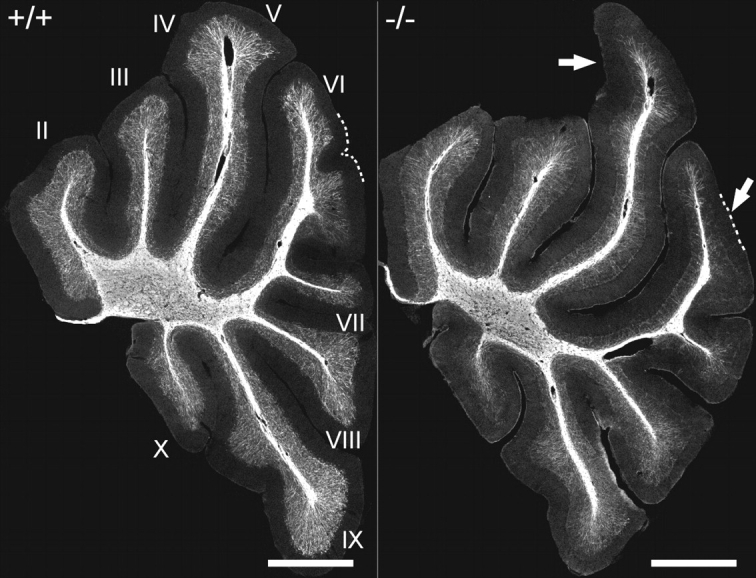
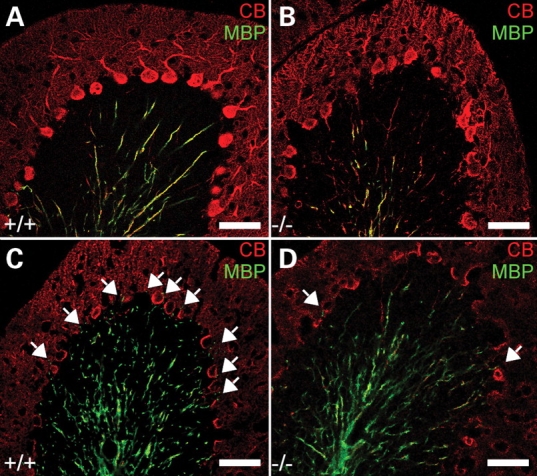
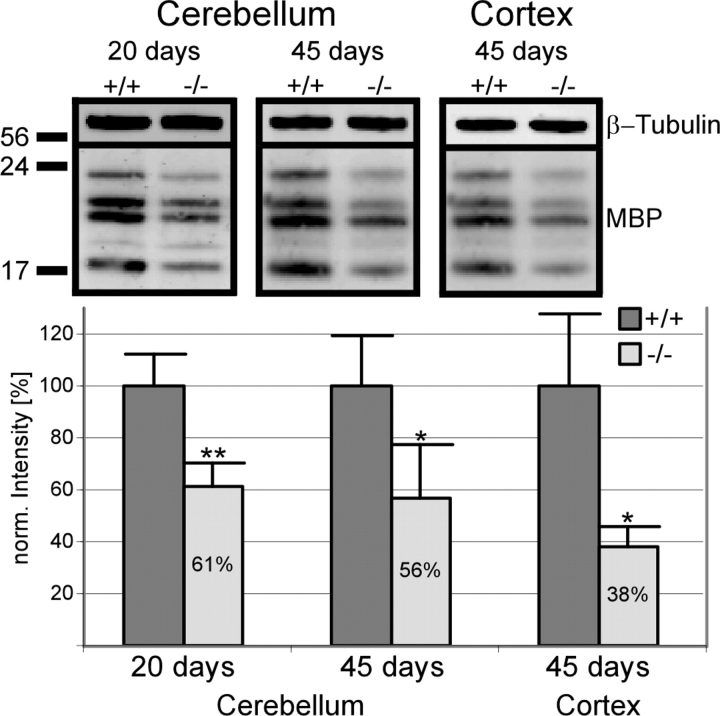
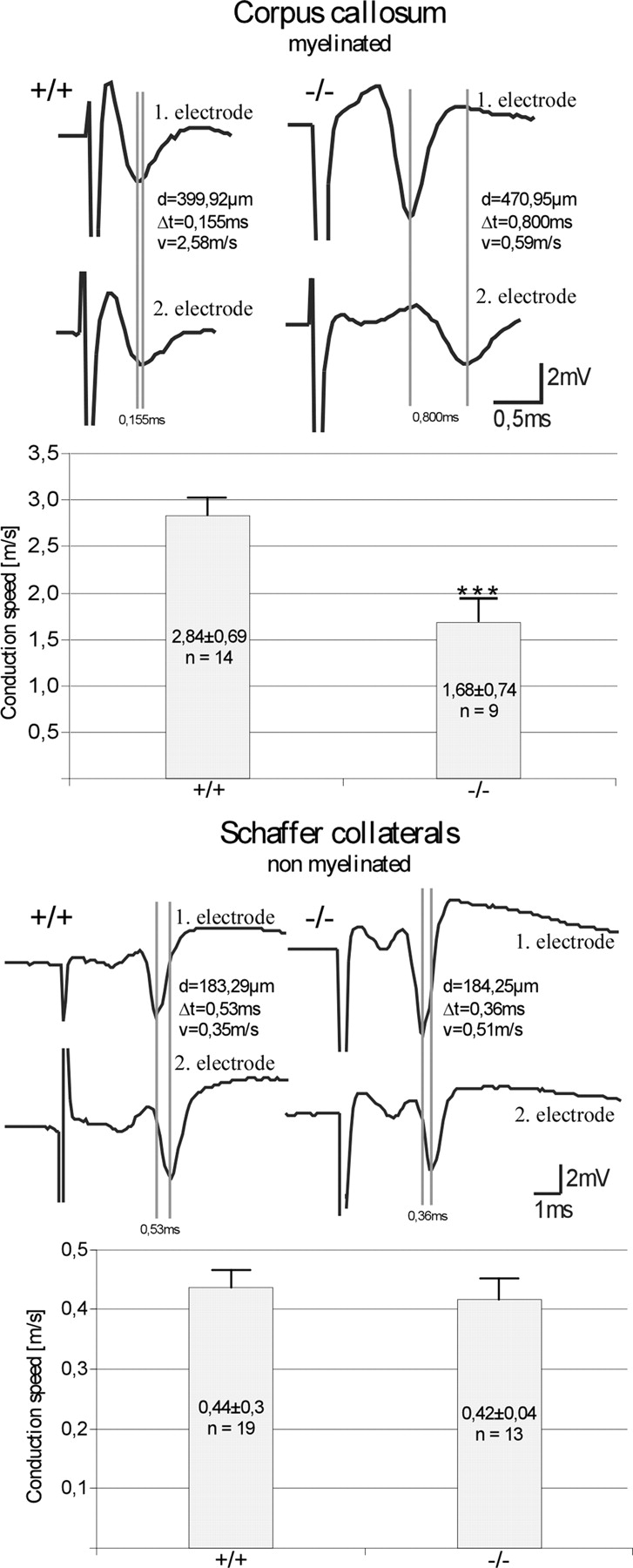
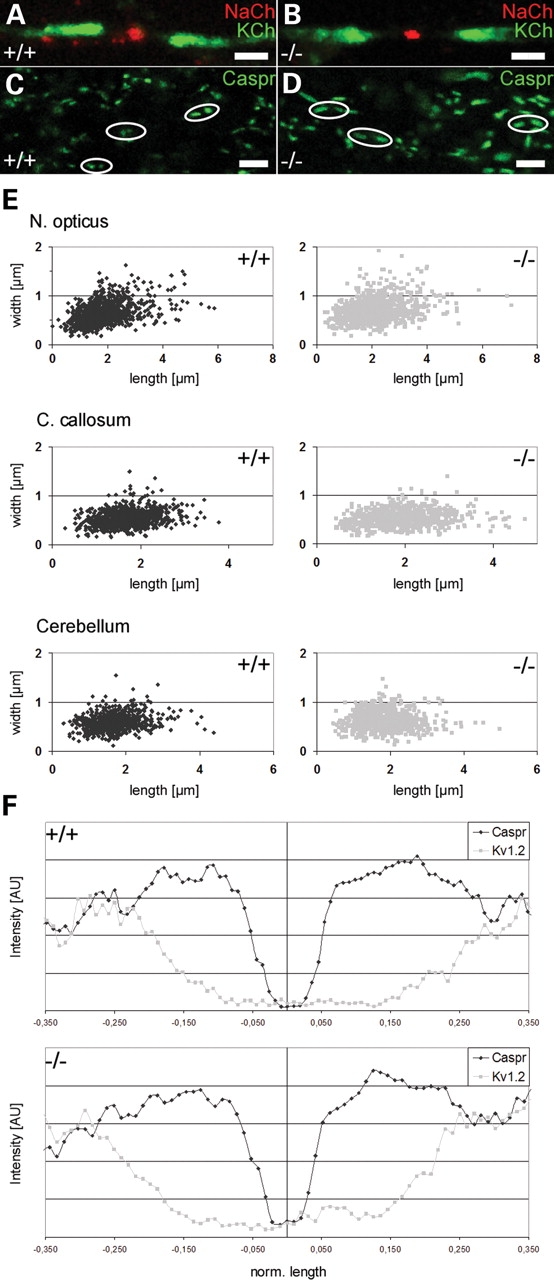
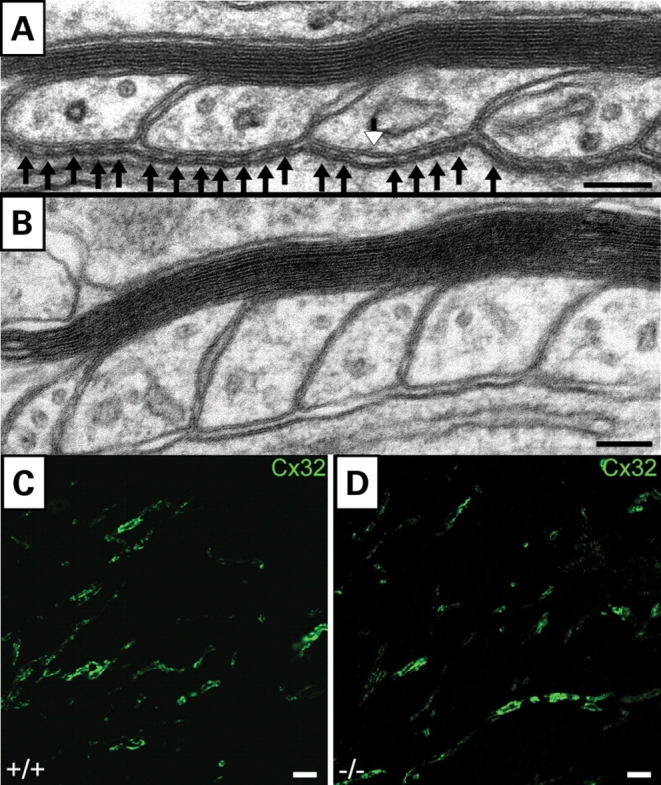
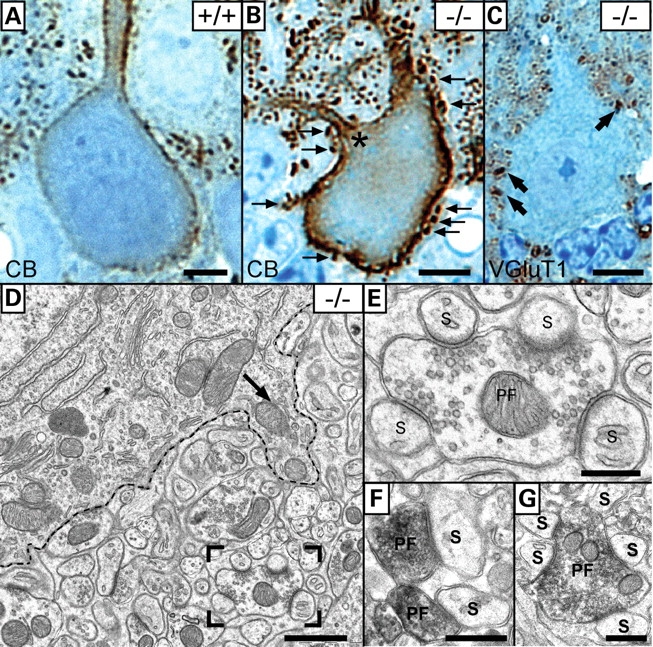
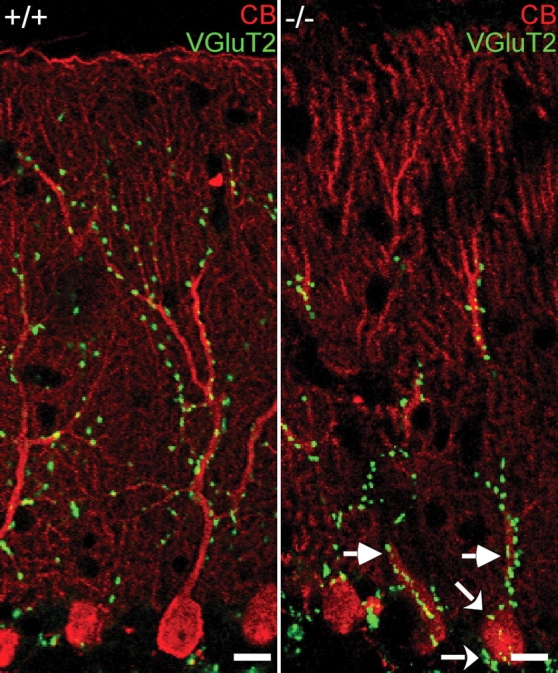
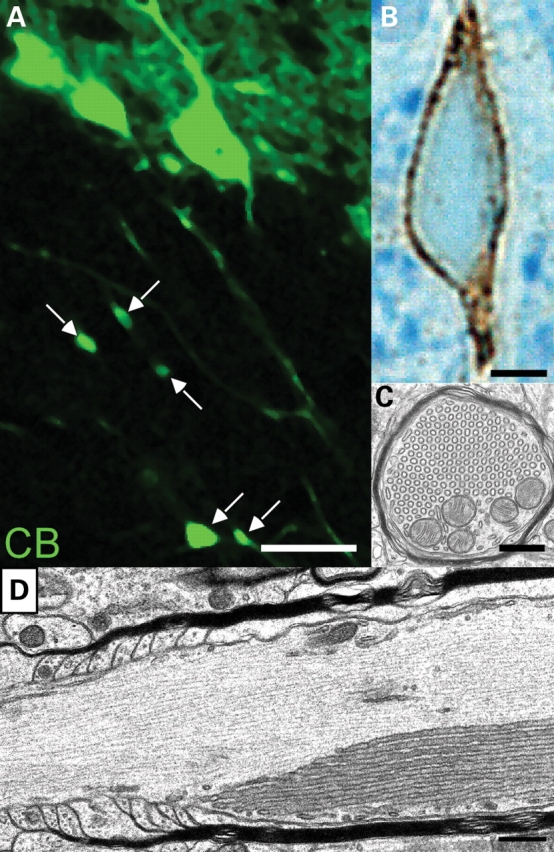
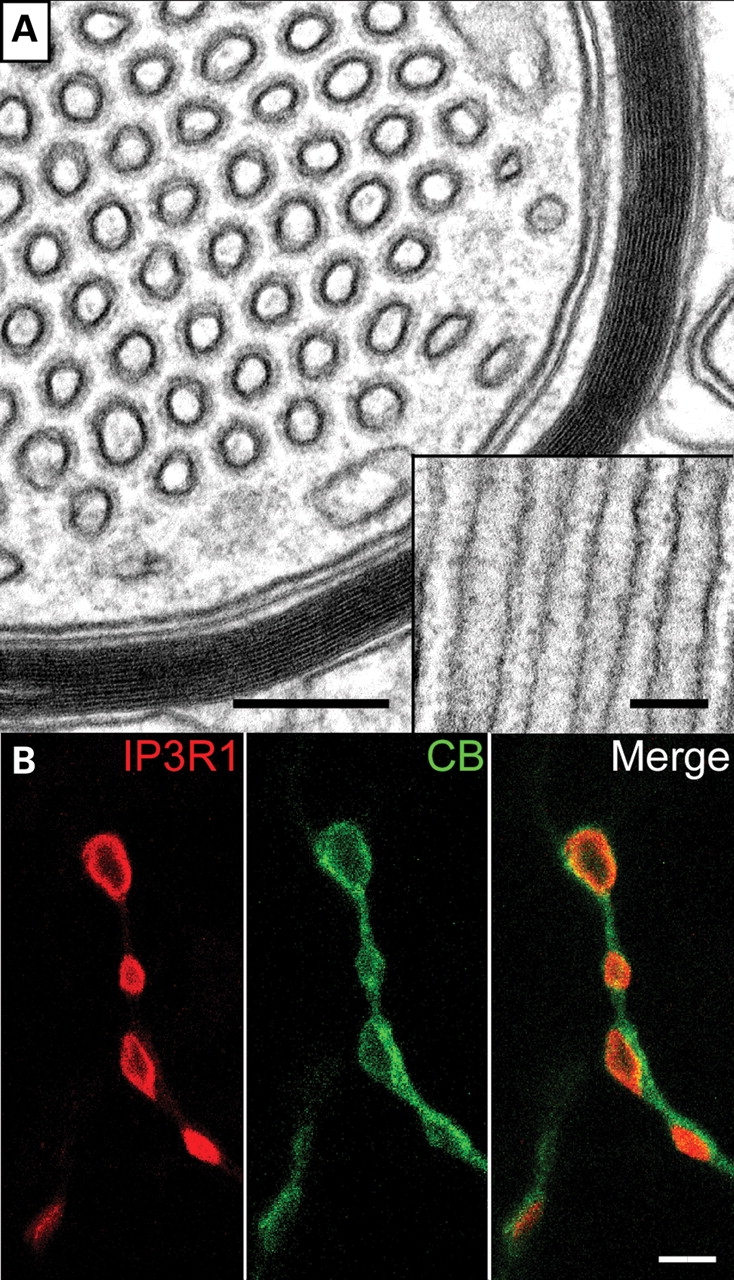
Ether lipids (ELs), particularly plasmalogens, are essential constituents of the mammalian central nervous system. The physiological role of ELs, in vivo, however is still enigmatic. In the present study, we characterized a mouse model carrying a targeted deletion of the peroxisomal dihydroxyacetonephosphate acyltransferase gene that results in the complete lack of ELs. Investigating the cerebellum of these mice, we observed: (i) defects in foliation patterning and delay in precursor granule cell migration, (ii) defects in myelination and concomitant reduction in the level of myelin basic protein, (iii) disturbances in paranode organization by extending the Caspr distribution and disrupting axo-glial septate-like junctions, (iv) impaired innervation of Purkinje cells by both parallel fibers and climbing fibers and (v) formation of axon swellings by the accumulation of inositol-tris-phosphate receptor 1 containing smooth ER-like tubuli. Functionally, conduction velocity of myelinated axons in the corpus callosum was significantly reduced. Most of these phenotypes were already apparent at P20 but still persisted in 1-year-old animals. In summary, these data show that EL deficiency results in severe developmental and lasting structural alterations at the cellular and network level of the cerebellum, and reveal an important role of ELs for proper brain function. Common molecular mechanisms that may underlie these phenotypes are discussed.
Figures
References
-
- Moser H.W. Molecular genetics of peroxisomal disorders. Front. Biosci. 2000;5:D298–D306. - PubMed
-
- Wanders R.J., Waterham H.R. Peroxisomal disorders I: biochemistry and genetics of peroxisome biogenesis disorders. Clin. Genet. 2005;67:107–133. - PubMed
-
- Gorgas K., Teigler A., Komljenovic D., Just W.W. The ether lipid-deficient mouse: tracking down plasmalogen functions. Biochim. Biophys. Acta. 2006;1763:1511–1526. - PubMed
-
- Nagan N., Zoeller R.A. Plasmalogens: biosynthesis and functions. Prog. Lipid Res. 2001;40:199–229. - PubMed
-
- Zoeller R.A., Morand O.H., Raetz C.R. A possible role for plasmalogens in protecting animal cells against photosensitized killing. J. Biol. Chem. 1988;263:11590–11596. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
