

Polyglutamine disruption of the huntingtin exon 1 N terminus triggers a complex aggregation mechanism
- PMID: 19270701
- PMCID: PMC2706102
- DOI: 10.1038/nsmb.1570
Polyglutamine disruption of the huntingtin exon 1 N terminus triggers a complex aggregation mechanism
Abstract
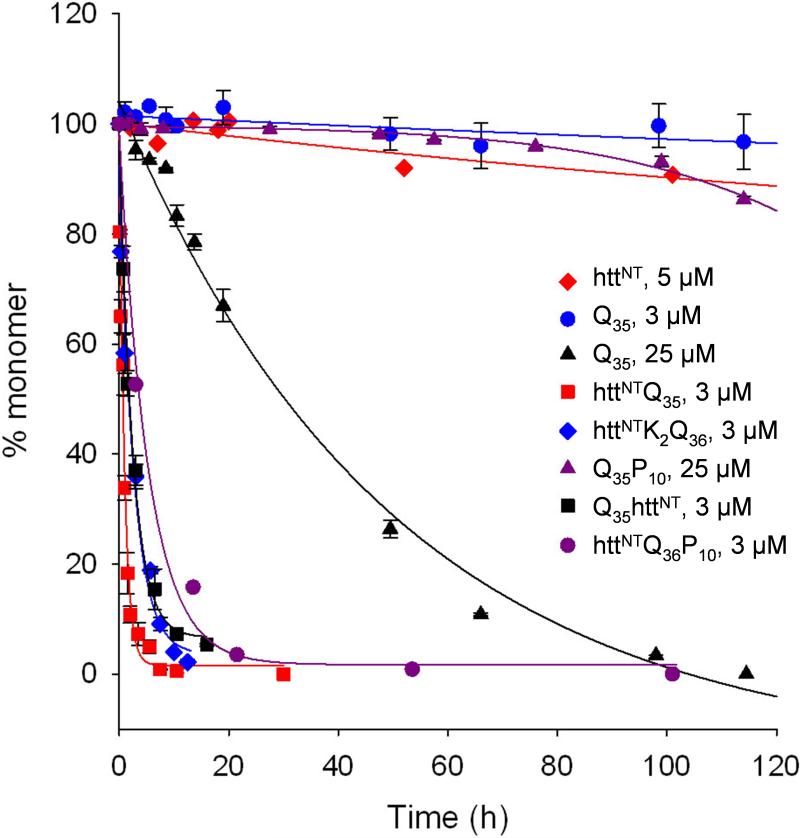
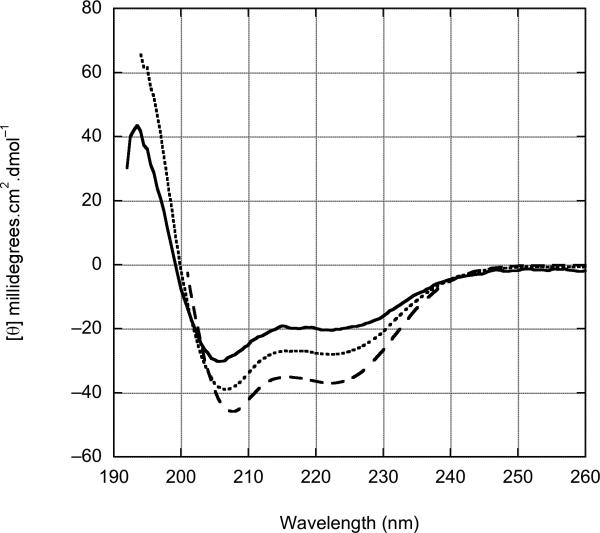
Simple polyglutamine (polyQ) peptides aggregate in vitro via a nucleated growth pathway directly yielding amyloid-like aggregates. We show here that the 17-amino-acid flanking sequence (HTT(NT)) N-terminal to the polyQ in the toxic huntingtin exon 1 fragment imparts onto this peptide a complex alternative aggregation mechanism. In isolation, the HTT(NT) peptide is a compact coil that resists aggregation. When polyQ is fused to this sequence, it induces in HTT(NT), in a repeat-length dependent fashion, a more extended conformation that greatly enhances its aggregation into globular oligomers with HTT(NT) cores and exposed polyQ. In a second step, a new, amyloid-like aggregate is formed with a core composed of both HTT(NT) and polyQ. The results indicate unprecedented complexity in how primary sequence controls aggregation within a substantially disordered peptide and have implications for the molecular mechanism of Huntington's disease.
Figures

















References
-
- Bates GP, Benn C. The polyglutamine diseases. In: Bates GP, Harper PS, Jones L, editors. Huntington's Disease. Oxford University Press; Oxford, U.K.: 2002. pp. 429–472.
-
- Bates GP, Harper PS, Jones L, editors. Huntington's Disease. Oxford University Press; Oxford, U.K.: 2002.
-
- Wetzel R. Misfolding and aggregation in Huntington's disease and other expanded polyglutamine repeat diseases. In: Dobson CM, Kelly JW, Ramirez-Alvarado M, editors. Protein Misfolding Diseases: Current and Emerging Principles and Therapies. Wiley; New York: 2009. in press.
-
- Arrasate M, Mitra S, Schweitzer ES, Segal MR, Finkbeiner S. Inclusion body formation reduces levels of mutant huntingtin and the risk of neuronal death. Nature. 2004;431:805–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
