

Morphological diversity and the roles of contingency, chance and determinism in african cichlid radiations
- PMID: 19270732
- PMCID: PMC2648897
- DOI: 10.1371/journal.pone.0004740
Morphological diversity and the roles of contingency, chance and determinism in african cichlid radiations
Abstract
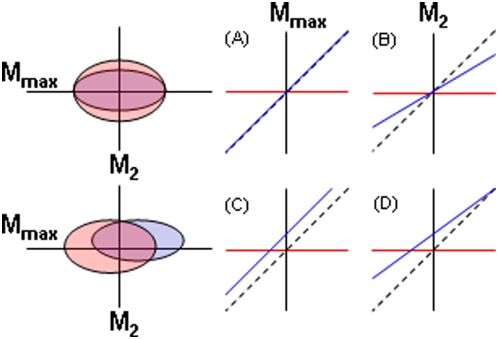
Background: Deterministic evolution, phylogenetic contingency and evolutionary chance each can influence patterns of morphological diversification during adaptive radiation. In comparative studies of replicate radiations, convergence in a common morphospace implicates determinism, whereas non-convergence suggests the importance of contingency or chance.
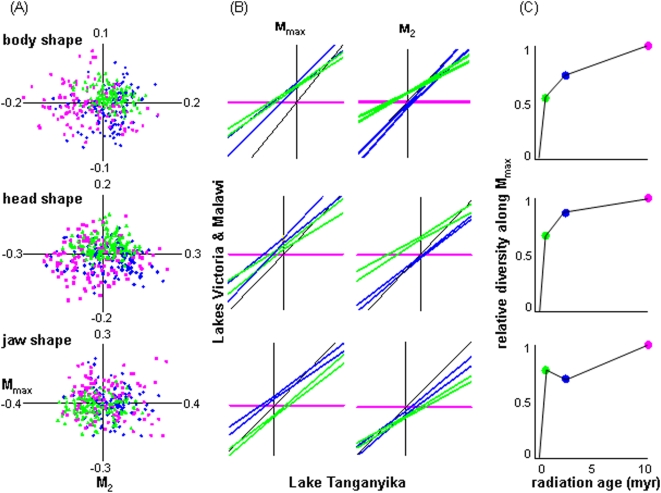
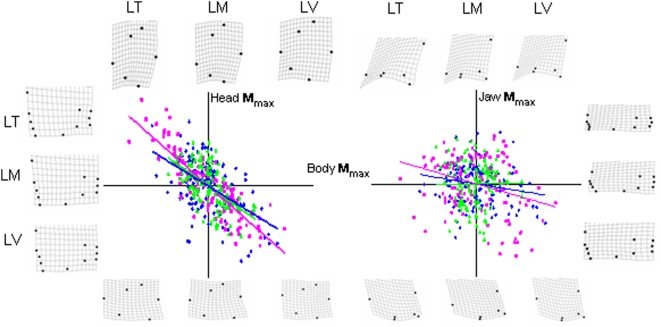
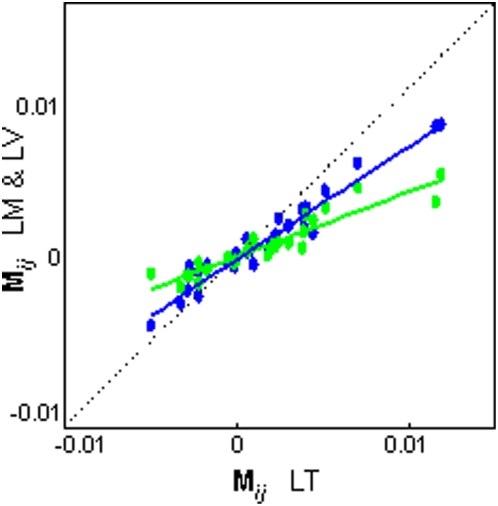
Methodology/principal findings: The endemic cichlid fish assemblages of the three African great lakes have evolved similar sets of ecomorphs but show evidence of non-convergence when compared in a common morphospace, suggesting the importance of contingency and/or chance. We then analyzed the morphological diversity of each assemblage independently and compared their axes of diversification in the unconstrained global morphospace. We find that despite differences in phylogenetic composition, invasion history, and ecological setting, the three assemblages are diversifying along parallel axes through morphospace and have nearly identical variance-covariance structures among morphological elements.
Conclusions/significance: By demonstrating that replicate adaptive radiations are diverging along parallel axes, we have shown that non-convergence in the common morphospace is associated with convergence in the global morphospace. Applying these complimentary analyses to future comparative studies will improve our understanding of the relationship between morphological convergence and non-convergence, and the roles of contingency, chance and determinism in driving morphological diversification.
Conflict of interest statement
Figures

References
-
- Schluter D. The ecology of adaptive radiation. Oxford: Oxford University Press.; 2000. p. 288.
-
- Schluter D. Ecological speciation in postglacial fishes. Phil Trans R Soc Ldn B. 1996;351:807–814.
-
- Losos JB, Jackman TR, Larson A, De Queiroz K, Rodriguez-Schettino L. Contingency and determinism in replicated adaptive radiation of island lizards. Science. 1998;279:2115–2118. - PubMed
-
- Rainey PB, Travisano M. Adaptive radiation in a heterogeneous environment. Nature. 1998;394:69–72. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
