

MMPs regulate both development and immunity in the tribolium model insect
- PMID: 19270735
- PMCID: PMC2649432
- DOI: 10.1371/journal.pone.0004751
MMPs regulate both development and immunity in the tribolium model insect
Abstract
Background: Matrix metalloproteinases (MMPs) are evolutionarily conserved and multifunctional effector molecules in development and homeostasis. In spite of previous, intensive investigation in vitro and in cell culture, their pleiotrophic functions in vivo are still not well understood.
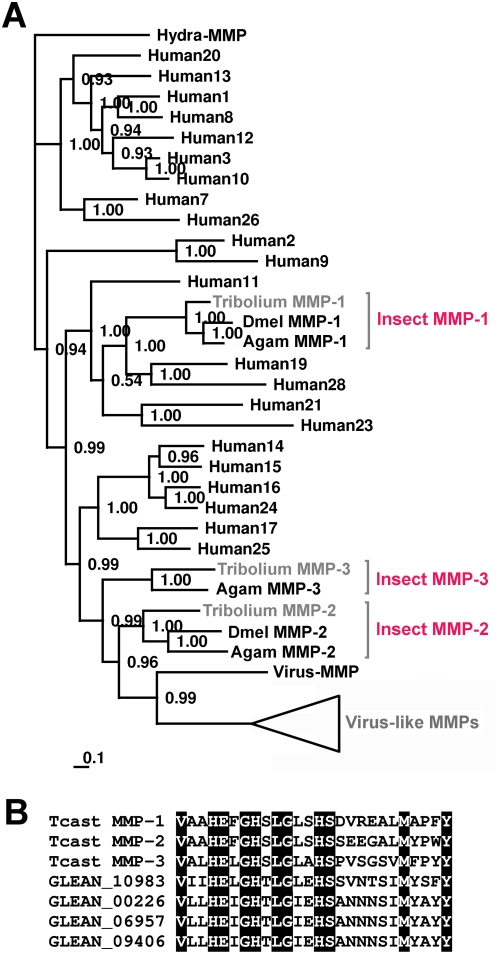
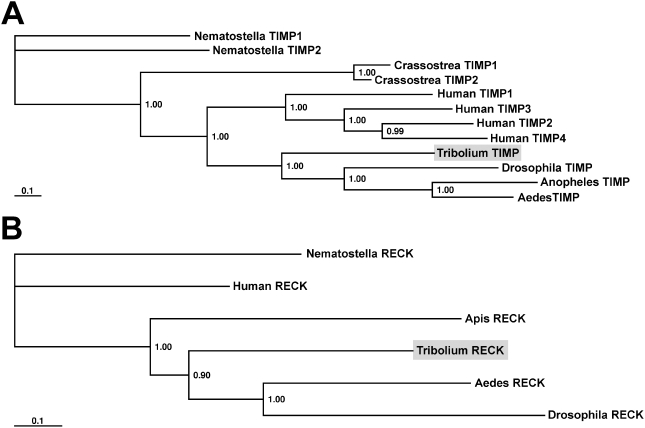
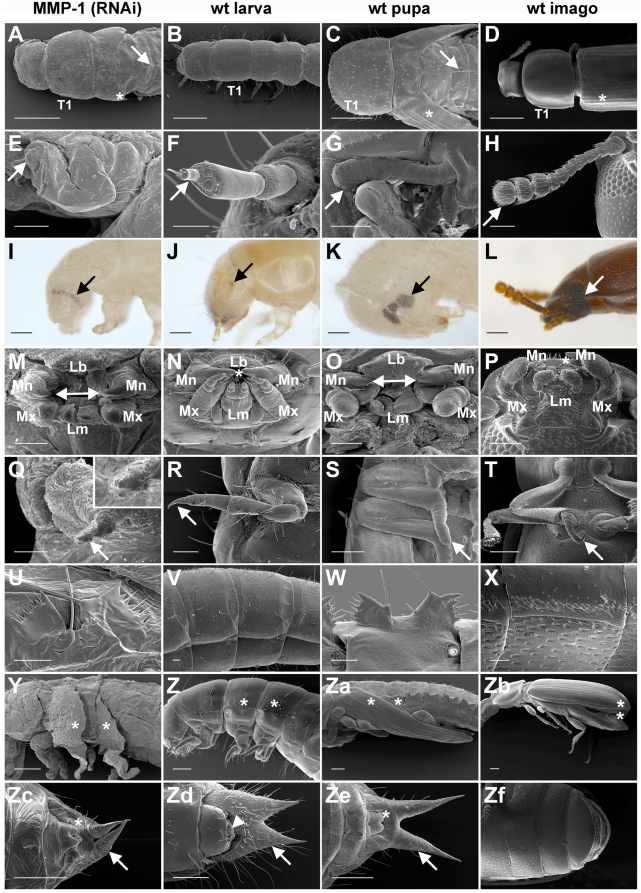
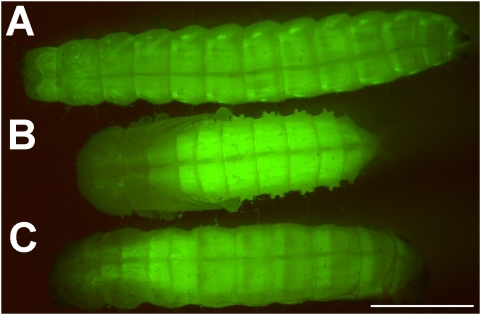
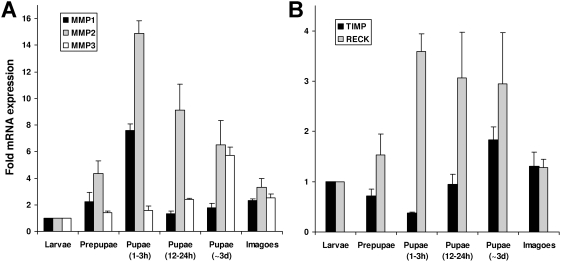
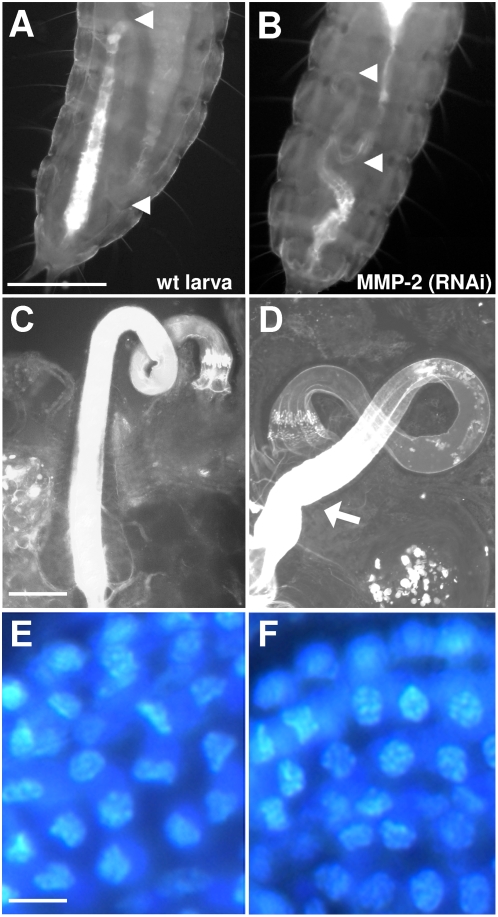
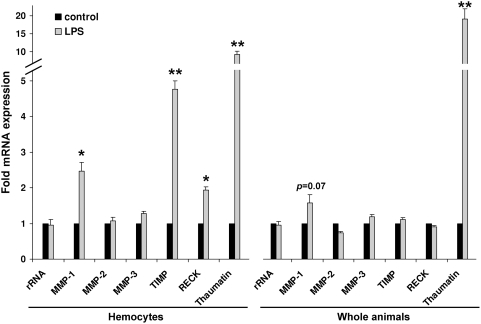
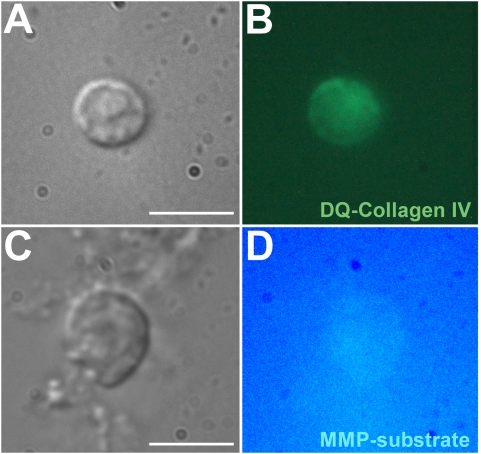
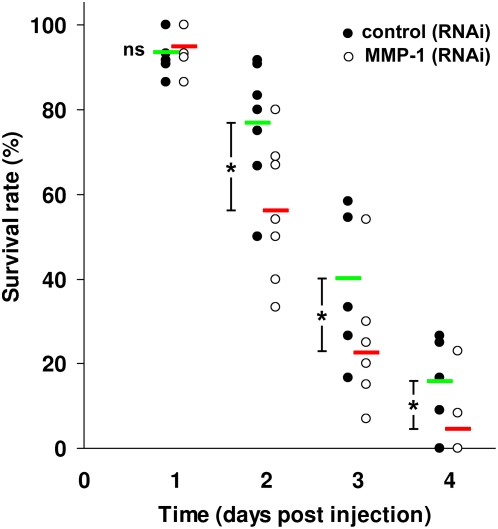
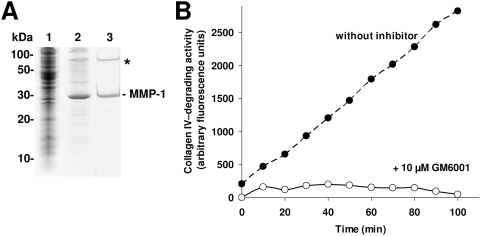
Methodology/principal findings: We show that the genetically amenable beetle Tribolium castaneum represents a feasible model organism to explore MMP functions in vivo. We silenced expression of three insect-type Tribolium MMP paralogs and their physiological inhibitors, TIMP and RECK, by dsRNA-mediated genetic interference (RNAi). Knock-down of MMP-1 arrested development during pupal morphogenesis giving phenotypes with altered antennae, compound eyes, wings, legs, and head. Parental RNAi-mediated knock-down of MMP-1 or MMP-2 resulted in larvae with non-lethal tracheal defects and with abnormal intestines, respectively, implicating additional roles of MMPs during beetle embryogenesis. This is different to findings from the fruit fly Drosophila melanogaster, in which MMPs have a negligible role in embryogenesis. Confirming pleiotrophic roles of MMPs our results also revealed that MMPs are required for proper insect innate immunity because systemic knock-down of Tribolium MMP-1 resulted in significantly higher susceptibility to the entomopathogenic fungus Beauveria bassiana. Moreover, mRNA levels of MMP-1, TIMP, and RECK, and also MMP enzymatic activity were significantly elevated in immune-competent hemocytes upon stimulation. To confirm collagenolytic activity of Tribolium MMP-1 we produced and purified recombinant enzyme and determined a similar collagen IV degrading activity as observed for the most related human MMP, MMP-19.
Conclusions/significance: This is the first study, to our knowledge, investigating the in vivo role of virtually all insect MMP paralogs along with their inhibitors TIMP and RECK in both insect development and immunity. Our results from the Tribolium model insect indicate that MMPs regulate tracheal and gut development during beetle embryogenesis, pupal morphogenesis, and innate immune defense reactions thereby revealing the evolutionarily conserved roles of MMPs.
Conflict of interest statement
Figures


References
-
- Nagase H, Woessner JF. Matrix metalloproteinases. J Biol Chem. 1999;274:21491–21494. - PubMed
-
- Vu TH, Werb Z. Matrix metalloproteinases: effectors of development and normal physiology. Genes Dev. 2000;14:2123–2133. - PubMed
-
- Bode W, Maskos K. Structural basis of the matrix metalloproteinases and their physiological inhibitors, the tissue inhibitors of metalloproteinases. Biol Chem. 2003;384:863–872. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
