

Silencing of PINK1 expression affects mitochondrial DNA and oxidative phosphorylation in dopaminergic cells
- PMID: 19270741
- PMCID: PMC2649444
- DOI: 10.1371/journal.pone.0004756
Silencing of PINK1 expression affects mitochondrial DNA and oxidative phosphorylation in dopaminergic cells
Abstract
Background: Mitochondrial dysfunction has been implicated in the pathogenesis of Parkinson's disease (PD). Impairment of the mitochondrial electron transport chain (ETC) and an increased frequency in deletions of mitochondrial DNA (mtDNA), which encodes some of the subunits of the ETC, have been reported in the substantia nigra of PD brains. The identification of mutations in the PINK1 gene, which cause an autosomal recessive form of PD, has supported mitochondrial involvement in PD. The PINK1 protein is a serine/threonine kinase localized in mitochondria and the cytosol. Its precise function is unknown, but it is involved in neuroprotection against a variety of stress signalling pathways.
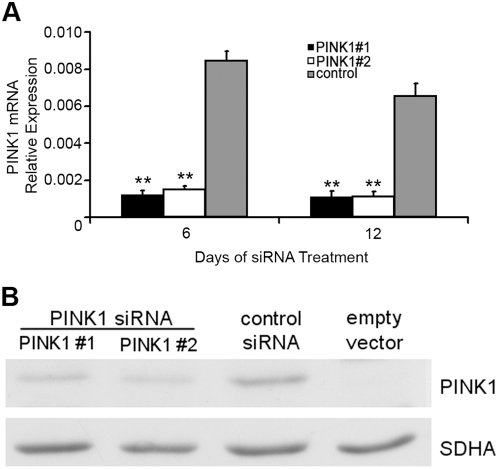
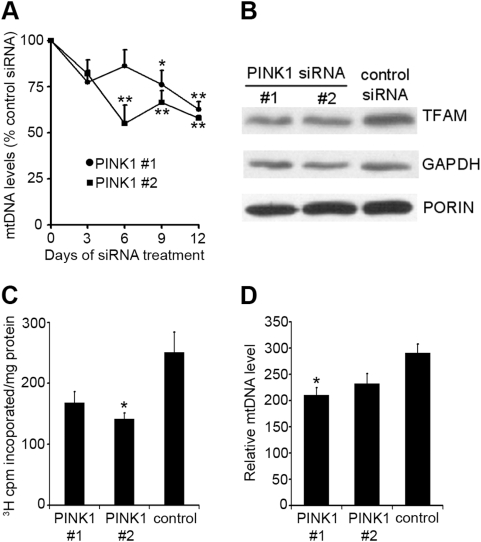
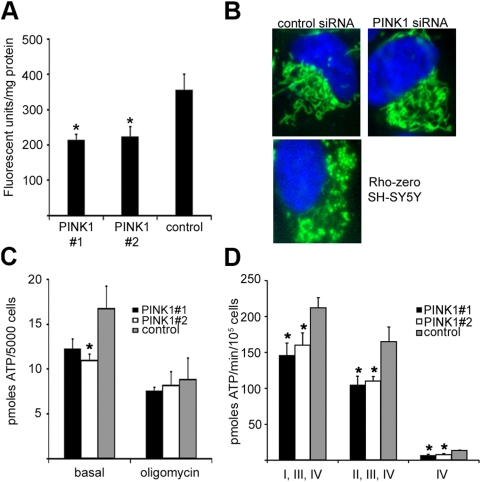
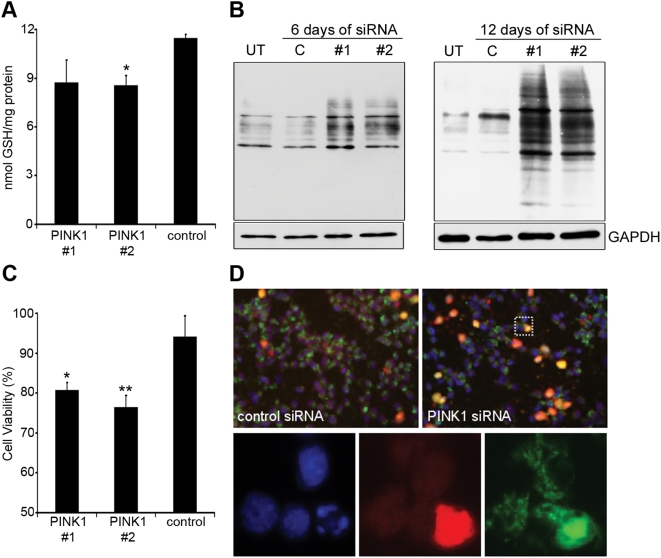
Methodology/principal findings: In this report we have investigated the effect of silencing PINK1 expression in human dopaminergic SH-SY5Y cells by siRNA on mtDNA synthesis and ETC function. Loss of PINK1 expression resulted in a decrease in mtDNA levels and mtDNA synthesis. We also report a concomitant loss of mitochondrial membrane potential and decreased mitochondrial ATP synthesis, with the activity of complex IV of the ETC most affected. This mitochondrial dysfunction resulted in increased markers of oxidative stress under basal conditions and increased cell death following treatment with the free radical generator paraquat.
Conclusions: This report highlights a novel function of PINK1 in mitochondrial biogenesis and a role in maintaining mitochondrial ETC activity. Dysfunction of both has been implicated in sporadic forms of PD suggesting that these may be key pathways in the development of the disease.
Conflict of interest statement
Figures
References
-
- Schapira AHV. Mitochondria in the aetiology and pathogenesis of Parkinson's disease. Lancet Neurol. 2008;7:97–109. - PubMed
-
- Schapira AH, Cooper JM, Dexter D, Jenner P, Clark JB, et al. Mitochondrial complex I deficiency in Parkinson's disease. Lancet. 1989;1 (8649):1269. - PubMed
-
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, et al. Chronic systemic pesticide exposure reproduces features of Parkinson's disease. Nat Neurosci. 2000;3:1301–1306. - PubMed
-
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Genetics. 2006;38:515–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
