

The chromatic input to cells of the magnocellular pathway of primates
- PMID: 19271925
- PMCID: PMC2810721
- DOI: 10.1167/9.2.15
The chromatic input to cells of the magnocellular pathway of primates
Abstract
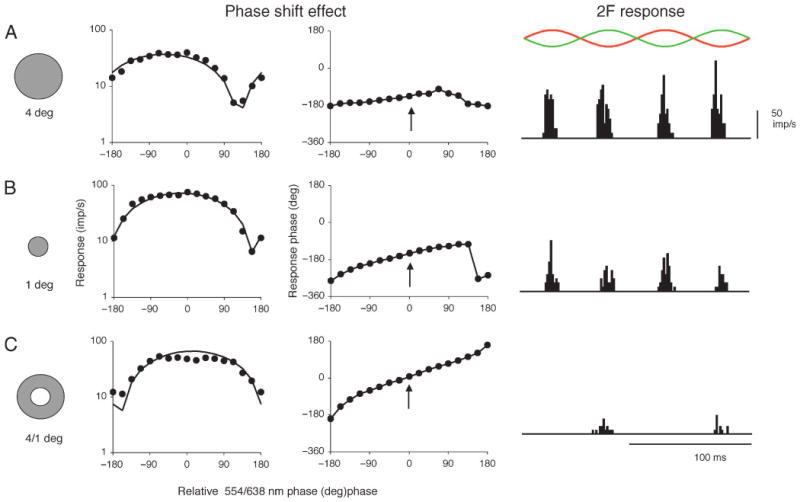
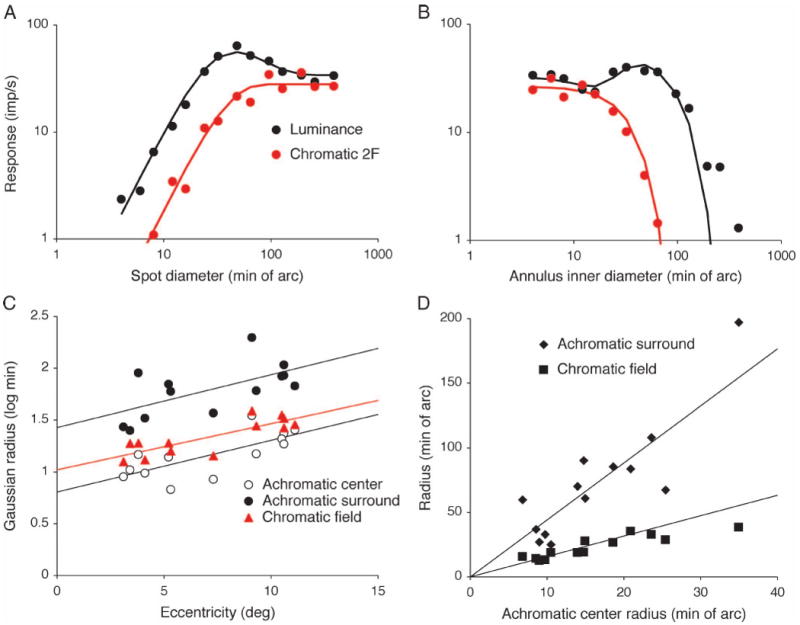
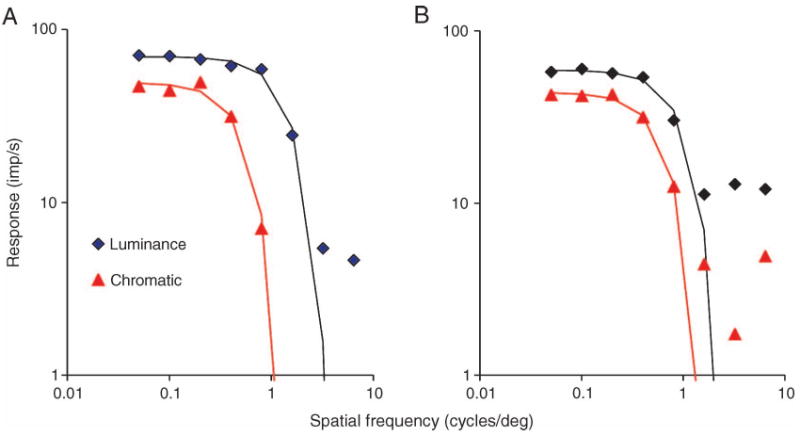
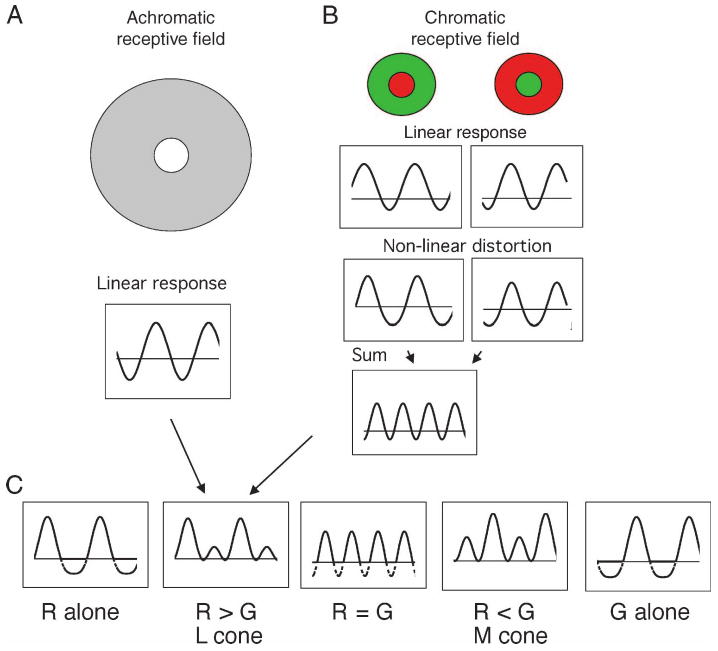
Parasol ganglion cells of the magnocellular (MC) pathway form the physiological substrate of a luminance channel underlying photometric tasks, but they also respond weakly to red-green chromatic modulation. This may take the form of a first-harmonic (1F) response to chromatic modulation at low temporal frequencies, and/or a second-harmonic (2F) response that is more marked at higher frequencies. It is shown here that both these responses originate from a receptive field component that is intermediate in size between center and surround, i.e., a discrete, chromatic receptive field is superimposed upon an achromatic center-surround structure. Its size is similar to the receptive field (center plus surround) of midget, parvocellular cells from the same retinal eccentricity. A 2F MC cell chromatic response component is shown to be present under cone silent substitution conditions, when only the middle- (M) or long-wavelength (L) cone is modulated. This and other features suggest it is a rectified response to a chromatic signal rather than a consequence of non-linear summation of M- and L-cone signals. A scheme is presented which could give rise to such responses. It is suggested that this chromatic input to MC cells can enhance motion signals to red-green borders close to equiluminance.
Figures







References
-
- Cavanagh P, Anstis S. The contribution of color to motion in normal and color-deficient observers. Vision Research. 1991;31:2109–2148. - PubMed
-
- Cavanagh P, Favreau OE. Color and luminance share a common motion pathway. Vision Research. 1985;25:1595–1601. - PubMed
-
- Chen Y, Martinez-Condez S, Macknik S, Swadlow H, Alonso JM, Lee B. Input to cells in macaque V1 revealed with a novel grating stimulus [Abstract] Journal of Vision. 2007. p. 57.p. 57a. http://journalofvisionorg/7/15/57/ - DOI
-
- Croner LJ, Kaplan E. Receptive fields of P and M ganglion cells across the primate retina. Vision Research. 1995;35:7–24. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
