

Review
doi: 10.2741/3328.
RNA-protein interactions in hepadnavirus reverse transcription
Affiliations
- PMID: 19273150
- PMCID: PMC3611959
- DOI: 10.2741/3328
Item in Clipboard
Review
RNA-protein interactions in hepadnavirus reverse transcription
Front Biosci (Landmark Ed).
.
Abstract
The small DNA genome of hepadnaviruses is replicated by reverse transcription via an RNA intermediate. This RNA "pregenome" contains important signals that control critical steps of viral replication, including RNA packaging, initiation of reverse transcription, and elongation of minus strand DNA, through specific interactions with the viral reverse transcriptase, the capsid protein, and host factors. In particular, the interaction between the viral reverse transcriptase and RNA pregenome requires a host chaperone complex composed of the heat shock protein 90 and its cochaperones.
Figures
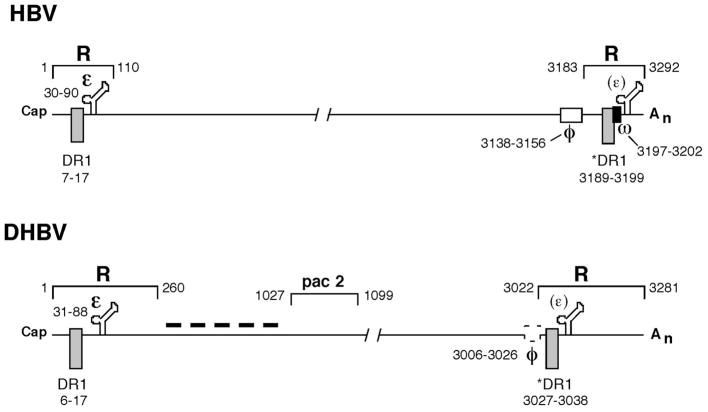
Summary of cis-acting signals on pgRNA of HBV and DHBV involved in RNA packaging and reverse transcription. The terminal redundancy (R) harbors the RNA packaging signal (ε), the DNA replication element direct repeat 1 (DR1), and the polyadenylation site. Note that the polyadenylation signal within the 5′ R is not utilized and omitted for clarity. The 3′ ε (in parenthesis) is not functional in mediating RNA packaging. The * symbol before the 3′ DR1 denotes the fact that only this copy of DR1 is used as the acceptor site during minus strand transfer. The nucleotide positions (with the cap site defined as nucleotide 1) of the various signals are indicated. Pac 2 denotes a second region of pgRNA required for RNA packaging in DHBV. The two elements required for specifying the acceptor site during minus strand DNA transfer, φ and ω, are indicated in HBV. The φ element proposed for DHBV (broken box) has not been experimentally confirmed. The dash line denotes the fact that the intervening sequences between ε and pac 2 also contribute to pgRNA packaging in DHBV. See text for details.

Proposed structures for the RNA packaging signal ε. The stem-loop structures for the HBV (A) and DHBV (B) ε RNA. The left scheme in each case represents the structures as originally proposed by Junker-Niepmann, et al (7) based on phylogenetic analysis and secondary structure prediction. The recently solved structures (by NMR) of the upper portion of the HBV and DHBV ε are shown to the right for comparison. Shaded boxes denote the apical loops as determined by NMR. The dash-lined box denotes the unstable base pairing in the middle of the upper stem of DHBV ε.
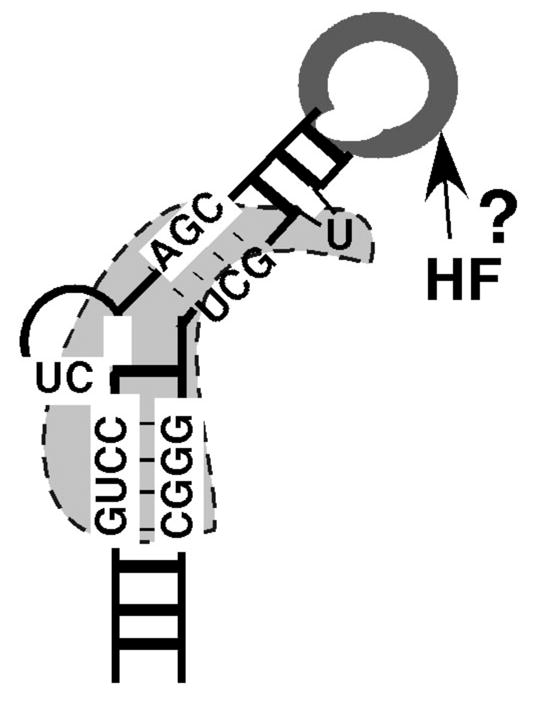
Model of HBV ε binding by RT and putative host factor. The HBV ε RNA is depicted as the typical stem loop structure. Critical sequences for RT binding (bounded by the dotted line) center around the internal bulge, including the first two nucleotides of the bulge, the upper portion of the lower stem and the lower portion of the upper stem. The apical loop, dispensable for RT binding but critical for pgRNA packaging, may interact with unknown host factor(s) (HF) in directing pgRNA packaging, and possibly protein priming. Adapted from Hu and Boyer (13). See text for detailed discussions of the model.
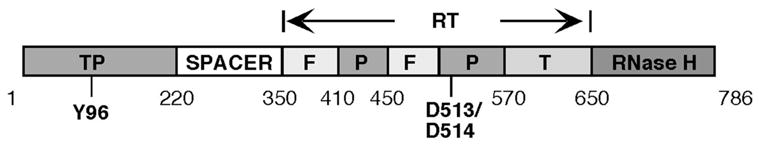
Domain structures of the hepadnavirus RT. The DHBV RT domains are shown schematically, with the approximate boundaries between the domains indicated. The central RT domain is further divided into the fingers (F), palm (P), and thumb (T) sub-domains through alignment with retroviral RT proteins. The primer tyrosine (Y96) in the TP domain and two catalytic aspartate residues (D513 and D514) in the highly conserved (among all RTs) YMDD motif of the palm sub-domain are indicated. Adapted from Wang et al (49).
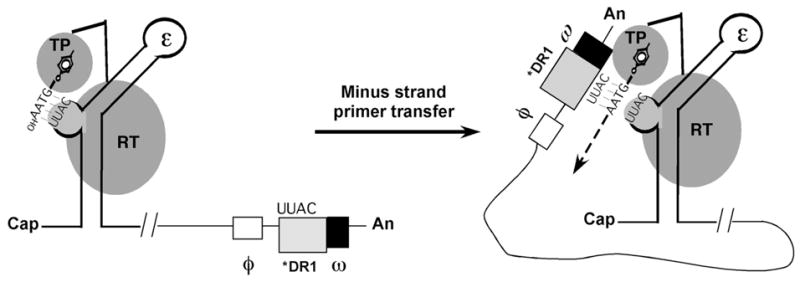
Protein priming and minus strand DNA transfer reactions in hepadnaviruses. Shown schematically are the ε stem-loop at the 5′ end of pgRNA and the RT protein with its TP and RT domains. (The RNase H domain is omitted for clarity.) Also shown are the RNA elements, φ, *DR1, and ω, which are involved in specifying the acceptor site on the 3′ end of pgRNA during minus strand DNA transfer following protein priming. Initiation of reverse transcription is triggered by the formation of the RNP complex between ε and RT. A specific tyrosine residue located at the TP domain acts as the primer for minus strand DNA synthesis. Using the 3′ 4 nucleotides of the ε bulge (UUAC in DHBV) as a template, RT synthesizes a short DNA oligomer (GTAA), which becomes covalently attached to RT via the primer tyrosine residue (protein priming). Subsequently, a template switch occurs whereby the nascent minus DNA-RT complex is translocated to the 3′ end of pgRNA. The nascent DNA strand then anneals to the homologous sequences at the acceptor site (*DR1) and minus strand DNA elongation continues. Long-range base-pairing interactions among sequences within ε, φ and ω may help to specify the acceptor site during minus strand DNA transfer. See text for details.
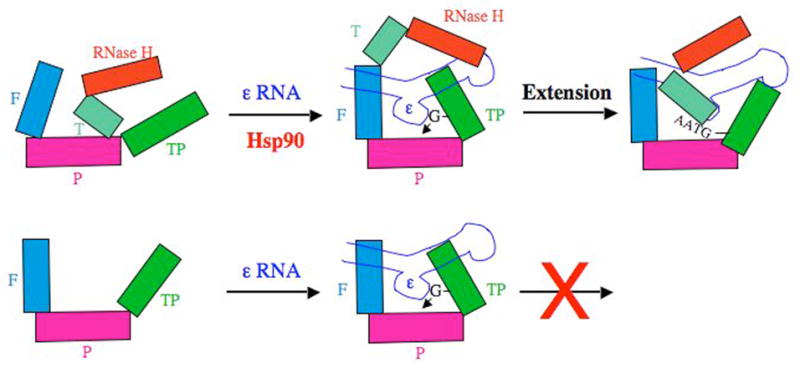
Working model for Hsp90-dependent and -independent folding of the DHBV RT and MiniRT2. The domains and subdomains of RT are depicted as blocks. Notations are as in Figure 4. The ε RNA is depicted as a stem-loop structure, with its internal bulge (the template for protein priming) facing the RT palm sub-domain. In the case of the full-length RT, the thumb sub-domain (and the RNase H domain) may prematurely interact with the palm sub-domain and preclude the TP domain from accessing the RT active site to establish a conformation competent for ε binding and initiation of protein priming. The Hsp90 chaperone complex is proposed to counteract this inhibitory effect of the thumb sub-domain and RNase H domain by preventing these inappropriate interactions and may, additionally, facilitate the productive interactions between the TP and the RT domains. In the case of MiniRT2, the removal of the thumb sub-domain and RNase H domain allows the mini RT to fold independently of Hsp90. On the other hand, following the initiation of protein priming (the covalent linkage of the dGMP residue to the TP domain), the thumb sub-domain has to access the RT active site (and the TP has to exit from it) in order to facilitate the subsequent DNA extension leading to the synthesis of the nascent DNA oligomer, GTAA. Lacking the thumb sub-domain, MiniRT2 is thus unable to carry out any DNA elongation. Adapted from Wang et al (49). See text for details.
Similar articles
-
Hepadnavirus reverse transcription initiates within the stem-loop of the RNA packaging signal and employs a novel strand transfer.J Virol. 1994 Jun;68(6):3536-43. doi: 10.1128/JVI.68.6.3536-3543.1994. J Virol. 1994. PMID: 8189492 Free PMC article.
-
Identification of a signal necessary for initiation of reverse transcription of the hepadnavirus genome.J Virol. 1991 Oct;65(10):5190-5. doi: 10.1128/JVI.65.10.5190-5195.1991. J Virol. 1991. PMID: 1895379 Free PMC article.
-
Why are hepadnaviruses DNA and not RNA viruses?Trends Microbiol. 1997 Nov;5(11):447-50. doi: 10.1016/s0966-842x(97)01141-4. Trends Microbiol. 1997. PMID: 9402701 Review.
-
Conservation of the HBV RNA element epsilon in nackednaviruses reveals ancient origin of protein-primed reverse transcription.Proc Natl Acad Sci U S A. 2021 Mar 30;118(13):e2022373118. doi: 10.1073/pnas.2022373118. Proc Natl Acad Sci U S A. 2021. PMID: 33753499 Free PMC article.
-
How do viral reverse transcriptases recognize their RNA genome?FEBS Lett. 1991 Aug 5;287(1-2):1-4. doi: 10.1016/0014-5793(91)80002-k. FEBS Lett. 1991. PMID: 1715279 Review.
Cited by
-
Inhibition of hepatitis B virus replication by the host zinc finger antiviral protein.PLoS Pathog. 2013;9(7):e1003494. doi: 10.1371/journal.ppat.1003494. Epub 2013 Jul 11. PLoS Pathog. 2013. PMID: 23853601 Free PMC article.
-
Drastic reduction in the production of subviral particles does not impair hepatitis B virus virion secretion.J Virol. 2009 Nov;83(21):11152-65. doi: 10.1128/JVI.00905-09. Epub 2009 Aug 12. J Virol. 2009. PMID: 19706705 Free PMC article.
-
In vitro epsilon RNA-dependent protein priming activity of human hepatitis B virus polymerase.J Virol. 2012 May;86(9):5134-50. doi: 10.1128/JVI.07137-11. Epub 2012 Feb 29. J Virol. 2012. PMID: 22379076 Free PMC article.
-
Protein-primed terminal transferase activity of hepatitis B virus polymerase.J Virol. 2013 Mar;87(5):2563-76. doi: 10.1128/JVI.02786-12. Epub 2012 Dec 19. J Virol. 2013. PMID: 23255788 Free PMC article.
-
RNA-Binding Motif Protein 24 (RBM24) Is Involved in Pregenomic RNA Packaging by Mediating Interaction between Hepatitis B Virus Polymerase and the Epsilon Element.J Virol. 2019 Mar 5;93(6):e02161-18. doi: 10.1128/JVI.02161-18. Print 2019 Mar 15. J Virol. 2019. PMID: 30626666 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
