

Role of the histone variant H2A.Z/Htz1p in TBP recruitment, chromatin dynamics, and regulated expression of oleate-responsive genes
- PMID: 19273605
- PMCID: PMC2668375
- DOI: 10.1128/MCB.01233-08
Role of the histone variant H2A.Z/Htz1p in TBP recruitment, chromatin dynamics, and regulated expression of oleate-responsive genes
Abstract
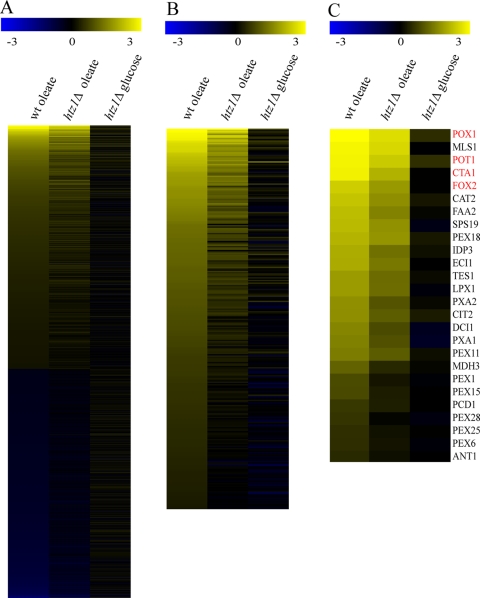
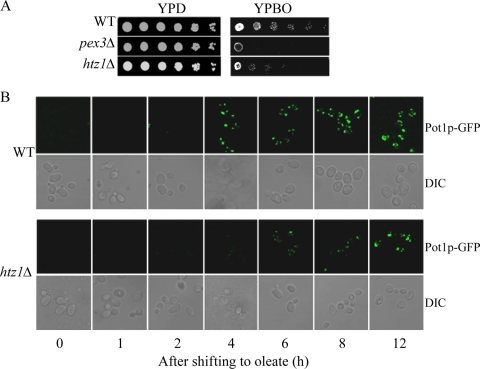
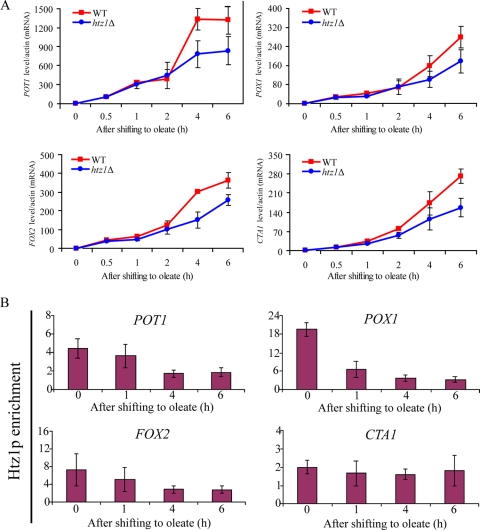
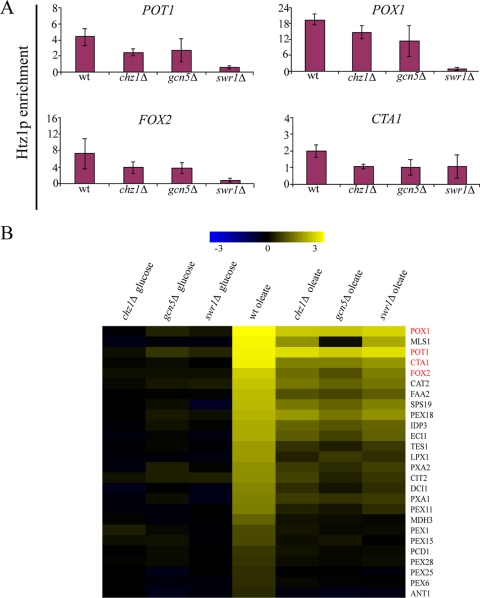
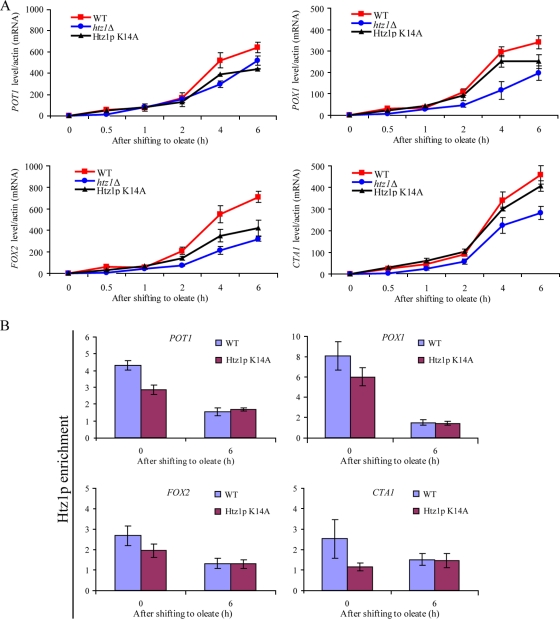
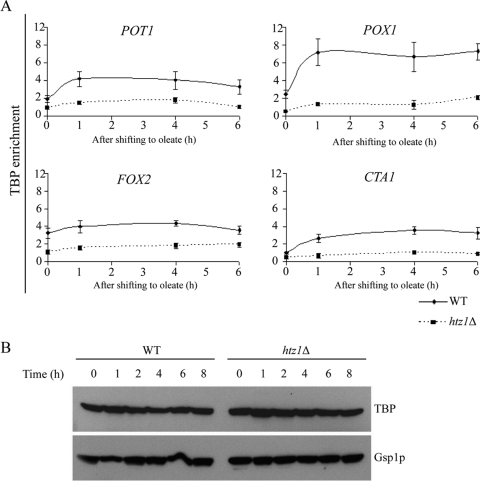
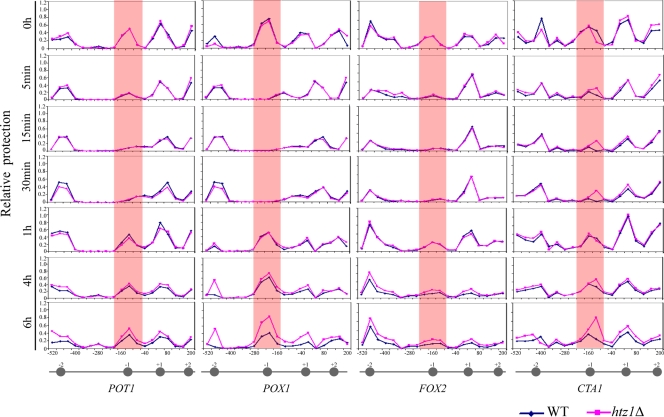
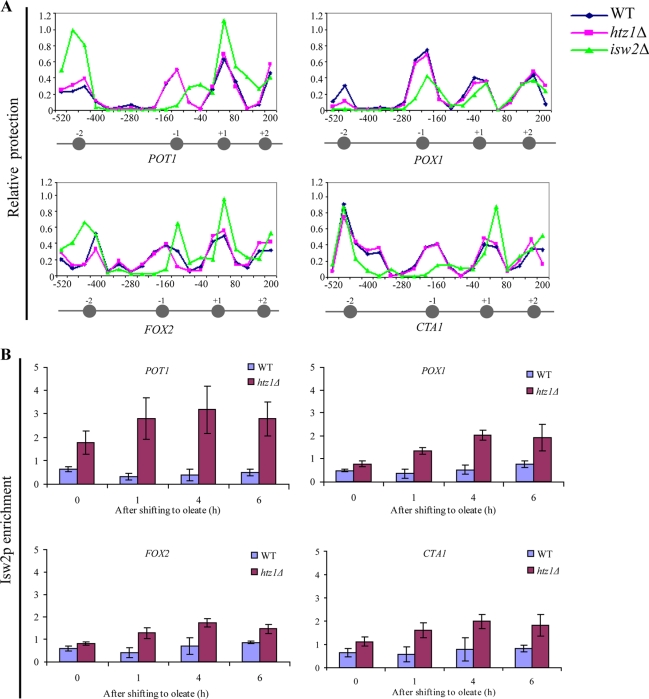
The histone variant H2A.Z (Htz1p) has been implicated in transcriptional regulation in numerous organisms, including Saccharomyces cerevisiae. Genome-wide transcriptome profiling and chromatin immunoprecipitation studies identified a role for Htz1p in the rapid and robust activation of many oleate-responsive genes encoding peroxisomal proteins, in particular POT1, POX1, FOX2, and CTA1. The Swr1p-, Gcn5p-, and Chz1p-dependent association of Htz1p with these promoters in their repressed states appears to establish an epigenetic marker for the rapid and strong expression of these highly inducible promoters. Isw2p also plays a role in establishing the nucleosome state of these promoters and associates stably in the absence of Htz1p. An analysis of the nucleosome dynamics and Htz1p association with these promoters suggests a complex mechanism in which Htz1p-containing nucleosomes at fatty acid-responsive promoters are disassembled upon initial exposure to oleic acid leading to the loss of Htz1p from the promoter. These nucleosomes reassemble at later stages of gene expression. While these new nucleosomes do not incorporate Htz1p, the initial presence of Htz1p appears to mark the promoter for sustained gene expression and the recruitment of TATA-binding protein.
Figures
References
-
- Aitchison, J. D., M. P. Rout, M. Marelli, G. Blobel, and R. W. Wozniak. 1995. Two novel related yeast nucleoporins Nup170p and Nup157p: complementation with the vertebrate homologue Nup155p and functional interactions with the yeast nuclear pore-membrane protein Pom152p. J. Cell Biol. 1311133-1148. - PMC - PubMed
-
- Albert, I., T. N. Mavrich, L. P. Tomsho, J. Qi, S. J. Zanton, S. C. Schuster, and B. F. Pugh. 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446572-576. - PubMed
-
- Bensinger, S. J., and P. Tontonoz. 2008. Integration of metabolism and inflammation by lipid-activated nuclear receptors. Nature 454470-477. - PubMed
-
- Berger, J., and D. E. Moller. 2002. The mechanisms of action of PPARs. Annu. Rev. Med. 53409-435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases