

Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation
- PMID: 19273624
- PMCID: PMC2699125
- DOI: 10.1084/jem.20081712
Circulating and gut-resident human Th17 cells express CD161 and promote intestinal inflammation
Abstract
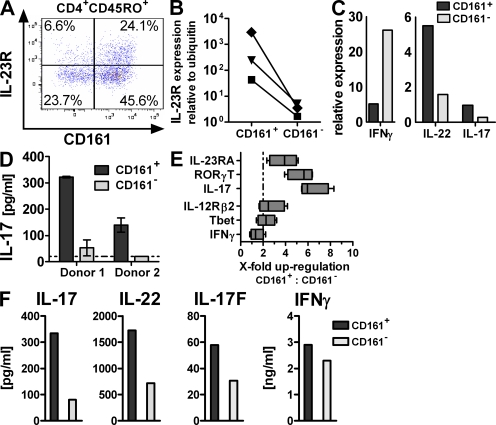
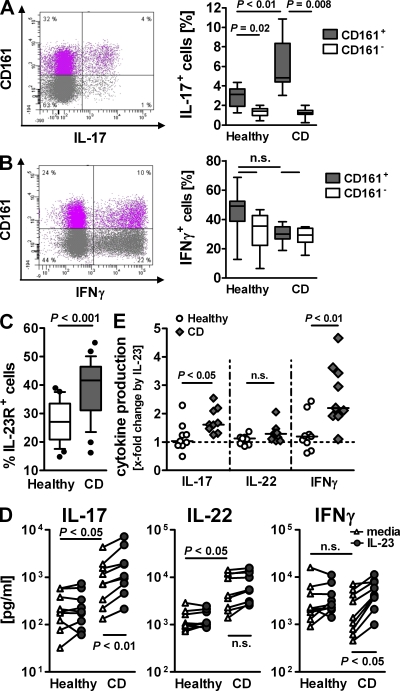
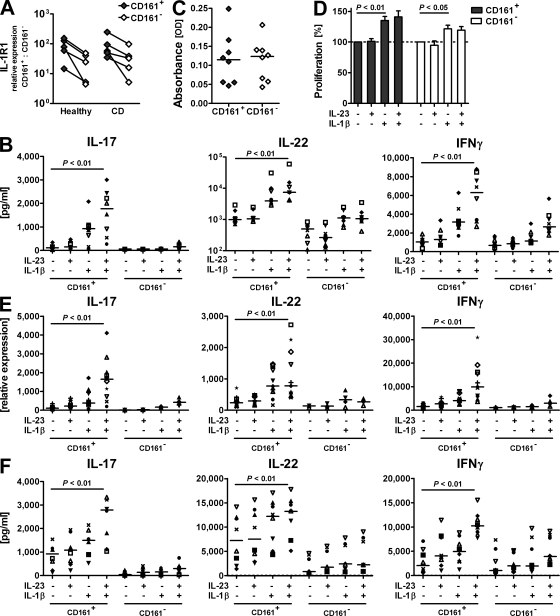
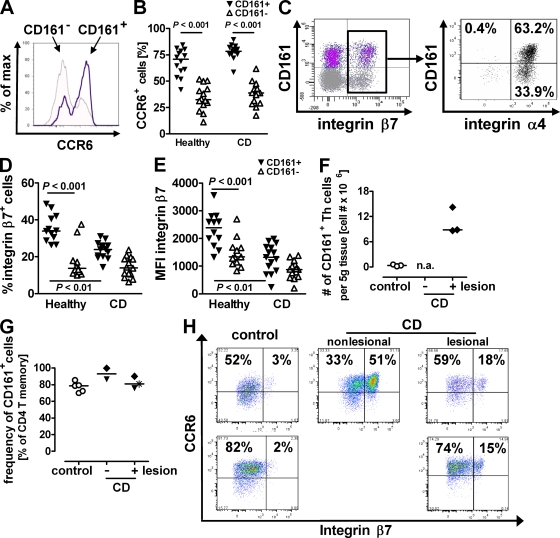
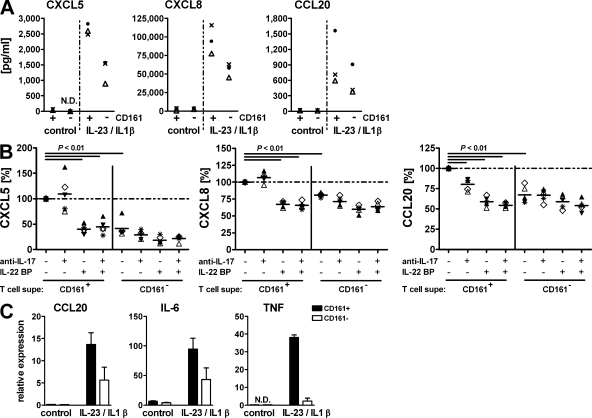
The C-type lectin-like receptor CD161, which has recently been described to promote T cell expansion, is expressed on a discrete subset of human CD4 T cells. The function of such cells, however, has remained elusive. We now demonstrate that CD161(+) CD4 T cells comprise a circulating and gut-resident T helper 17 (Th17) cell population. During Crohn's disease (CD), these CD161(+) cells display an activated Th17 phenotype, as indicated by increased expression of interleukin (IL)-17, IL-22, and IL-23 receptor. CD161(+) CD4 T cells from CD patients readily produce IL-17 and interferon gamma upon stimulation with IL-23, whereas, in healthy subjects, priming by additional inflammatory stimuli such as IL-1beta was required to enable IL-23-induced cytokine release. Circulating CD161(+) Th17 cells are imprinted for gut homing, as indicated by high levels of CC chemokine receptor 6 and integrin beta7 expression. Supporting their colitogenic phenotype, CD161(+) Th17 cells were found in increased numbers in the inflammatory infiltrate of CD lesions and induced expression of inflammatory mediators by intestinal cells. Our data identify CD161(+) CD4 T cells as a resting Th17 pool that can be activated by IL-23 and mediate destructive tissue inflammation.
Figures
References
-
- Kastelein R.A., Hunter C.A., Cua D.J. 2007. Discovery and biology of IL-23 and IL-27: related but functionally distinct regulators of inflammation.Annu. Rev. Immunol. 25:221–242 - PubMed
-
- McGeachy M.J., Cua D.J. 2008. Th17 cell differentiation: the long and winding road.Immunity. 28:445–453 - PubMed
-
- Cho J.H. 2008. The genetics and immunopathogenesis of inflammatory bowel disease.Nat. Rev. Immunol. 8:458–466 - PubMed
-
- Parham C., Chirica M., Timans J., Vaisberg E., Travis M., Cheung J., Pflanz S., Zhang R., Singh K.P., Vega F., et al. 2002. A receptor for the heterodimeric cytokine IL-23 is composed of IL-12Rbeta1 and a novel cytokine receptor subunit, IL-23R.J. Immunol. 168:5699–5708 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
