

Post-transcriptional regulation of cadherin-11 expression by GSK-3 and beta-catenin in prostate and breast cancer cells
- PMID: 19274078
- PMCID: PMC2650783
- DOI: 10.1371/journal.pone.0004797
Post-transcriptional regulation of cadherin-11 expression by GSK-3 and beta-catenin in prostate and breast cancer cells
Abstract
Background: The cell-cell adhesion molecule cadherin-11 is important in embryogenesis and bone morphogenesis, invasion of cancer cells, lymphangiogenesis, homing of cancer cells to bone, and rheumatoid arthritis. However, very little is known about the regulation of cadherin-11 expression.
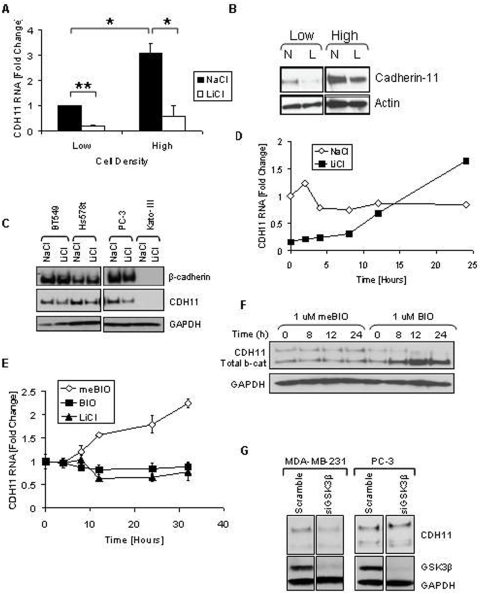
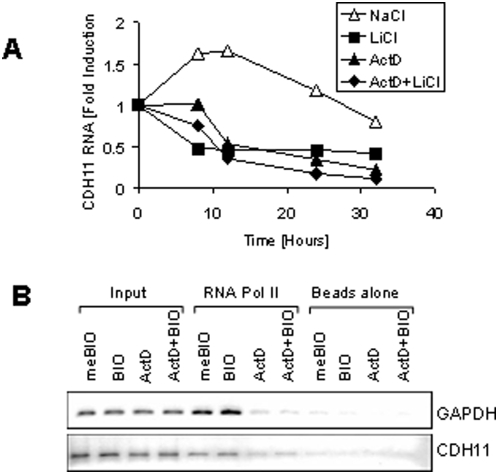
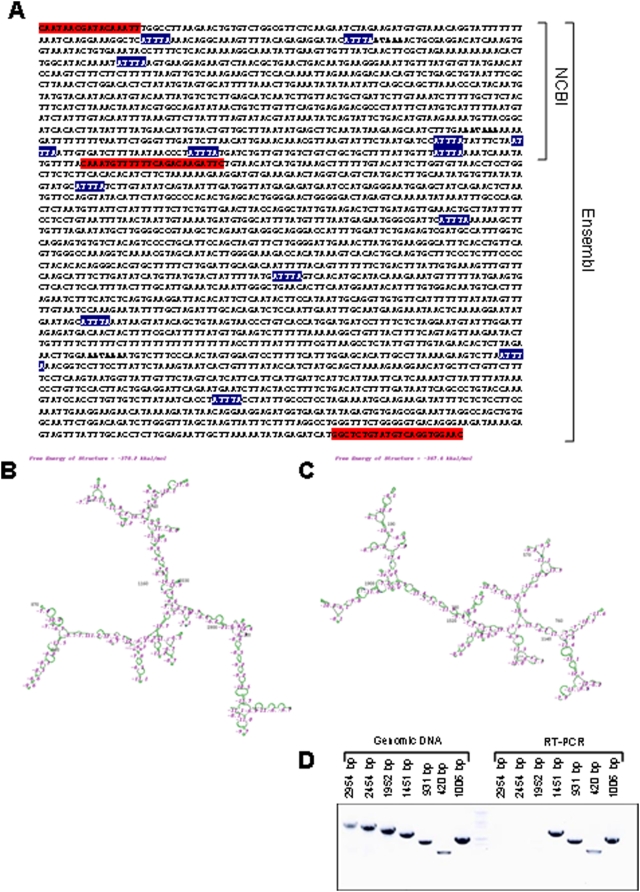
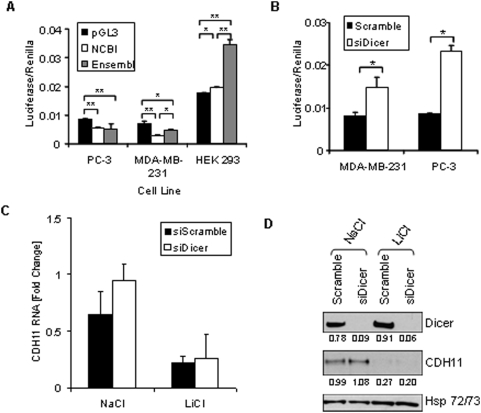
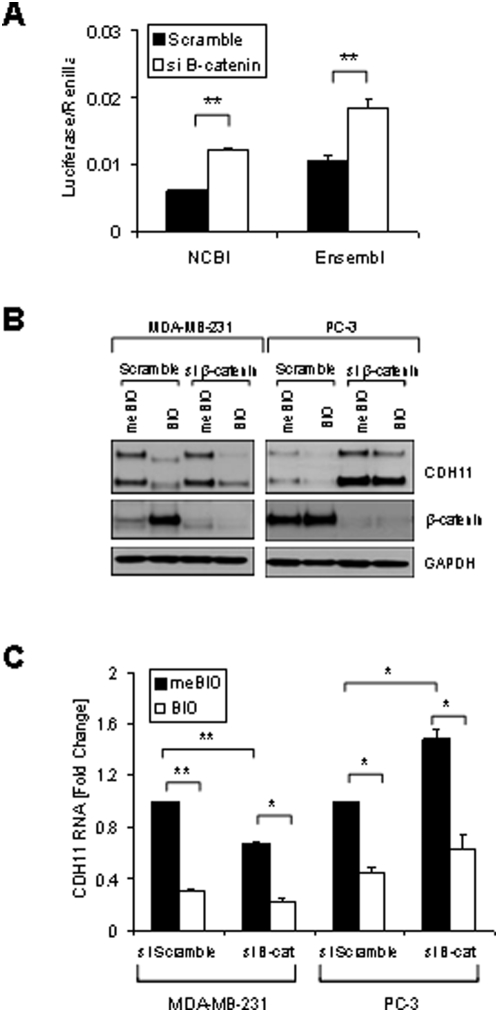
Methodology/principal findings: Here we show that cell density and GSK-3beta regulate cadherin-11 levels in cancer cells. Inactivation of GSK3beta with lithium chloride or the GSK3 inhibitor BIO and GSK3beta knockdown with siRNA repressed cadherin-11 mRNA and protein levels. RNA Polymerase II chromatin immunoprecipitation experiments showed that inhibition of GSK3 does not affect cadherin-11 gene transcription. Although the cadherin-11 3'UTR contains putative microRNA target sites and is regulated by Dicer, its stability is not regulated by GSK3 inhibition or density. Our data show that GSK3beta regulates cadherin-11 expression in two ways: first a beta-catenin-independent regulation of cadherin-11 steady state mRNA levels, and second a beta-catenin-dependent effect on cadherin-11 3'UTR stability and protein translation.
Conclusions: Cadherin-11 mRNA and protein levels are regulated by the activity of GSK3beta and a significant degree of this regulation is exerted by the GSK3 target, beta-catenin, at the level of the cadherin-11 3'UTR.
Conflict of interest statement
Figures
References
-
- Simonneau L, Kitagawa M, Suzuki S, Thiery JP. Cadherin 11 expression marks the mesenchymal phenotype: towards new functions for cadherins? Cell Adhes Commun. 1995;3:115–130. - PubMed
-
- Lee DM, Kiener HP, Agarwal SK, Noss EH, Watts GF, et al. Cadherin-11 in synovial lining formation and pathology in arthritis. Science. 2007;315:1006–1010. - PubMed
-
- Bussemakers MJ, Van Bokhoven A, Tomita K, Jansen CF, Schalken JA. Complex cadherin expression in human prostate cancer cells. Int J Cancer. 2000;85:446–450. - PubMed
-
- Guise TA. Molecular mechanisms of osteolytic bone metastases. Cancer. 2000;88:2892–2898. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
