

The cholinesterase-like domain, essential in thyroglobulin trafficking for thyroid hormone synthesis, is required for protein dimerization
- PMID: 19276074
- PMCID: PMC2676005
- DOI: 10.1074/jbc.M806898200
The cholinesterase-like domain, essential in thyroglobulin trafficking for thyroid hormone synthesis, is required for protein dimerization
Abstract
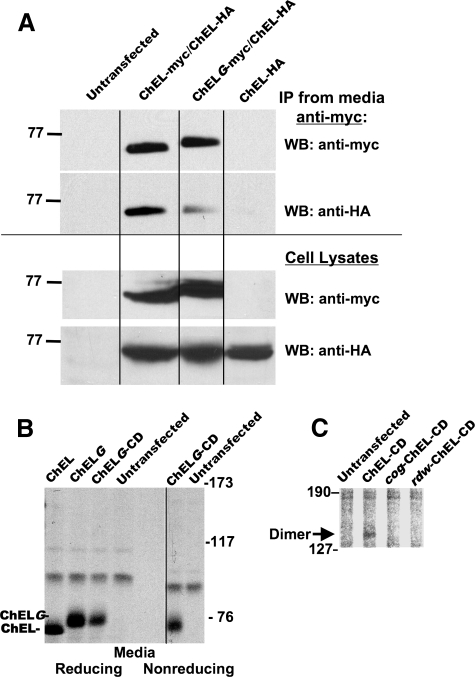
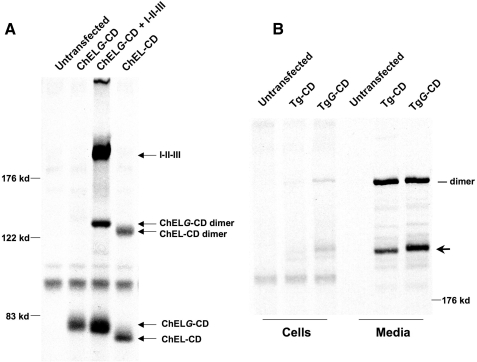
The carboxyl-terminal cholinesterase-like (ChEL) domain of thyroglobulin (Tg) has been identified as critically important in Tg export from the endoplasmic reticulum. In a number of human kindreds suffering from congenital hypothyroidism, and in the cog congenital goiter mouse and rdw rat dwarf models, thyroid hormone synthesis is inhibited because of mutations in the ChEL domain that block protein export from the endoplasmic reticulum. We hypothesize that Tg forms homodimers through noncovalent interactions involving two predicted alpha-helices in each ChEL domain that are homologous to the dimerization helices of acetylcholinesterase. This has been explored through selective epitope tagging of dimerization partners and by inserting an extra, unpaired Cys residue to create an opportunity for intermolecular disulfide pairing. We show that the ChEL domain is necessary and sufficient for Tg dimerization; specifically, the isolated ChEL domain can dimerize with full-length Tg or with itself. Insertion of an N-linked glycan into the putative upstream dimerization helix inhibits homodimerization of the isolated ChEL domain. However, interestingly, co-expression of upstream Tg domains, either in cis or in trans, overrides the dimerization defect of such a mutant. Thus, although the ChEL domain provides a nidus for Tg dimerization, interactions of upstream Tg regions with the ChEL domain actively stabilizes the Tg dimer complex for intracellular transport.
Figures






Similar articles
-
Cis and trans actions of the cholinesterase-like domain within the thyroglobulin dimer.J Biol Chem. 2010 Jun 4;285(23):17564-73. doi: 10.1074/jbc.M110.111641. Epub 2010 Mar 30. J Biol Chem. 2010. PMID: 20353937 Free PMC article.
-
The acetylcholinesterase homology region is essential for normal conformational maturation and secretion of thyroglobulin.J Biol Chem. 2004 Apr 23;279(17):17085-9. doi: 10.1074/jbc.M314042200. Epub 2004 Feb 5. J Biol Chem. 2004. PMID: 14764582
-
The cholinesterase-like domain of thyroglobulin functions as an intramolecular chaperone.J Clin Invest. 2008 Aug;118(8):2950-8. doi: 10.1172/JCI35164. J Clin Invest. 2008. PMID: 18596923 Free PMC article.
-
Thyroglobulin From Molecular and Cellular Biology to Clinical Endocrinology.Endocr Rev. 2016 Feb;37(1):2-36. doi: 10.1210/er.2015-1090. Epub 2015 Nov 23. Endocr Rev. 2016. PMID: 26595189 Free PMC article. Review.
-
The importance of thyroglobulin structure for thyroid hormone biosynthesis.Biochimie. 1999 May;81(5):505-9. doi: 10.1016/s0300-9084(99)80102-3. Biochimie. 1999. PMID: 10403182 Review.
Cited by
-
Thyroid hormone synthesis continues despite biallelic thyroglobulin mutation with cell death.JCI Insight. 2021 Jun 8;6(11):e148496. doi: 10.1172/jci.insight.148496. JCI Insight. 2021. PMID: 33914707 Free PMC article.
-
Novel TG-FGFR1 and TRIM33-NTRK1 transcript fusions in papillary thyroid carcinoma.Genes Chromosomes Cancer. 2019 Aug;58(8):558-566. doi: 10.1002/gcc.22737. Epub 2019 Feb 18. Genes Chromosomes Cancer. 2019. PMID: 30664823 Free PMC article.
-
Molecular assembly of thyroglobulin induced by in vitro nitric oxide treatments: implication its role in thyroid cells.Protein J. 2013 Dec;32(8):619-25. doi: 10.1007/s10930-013-9524-z. Protein J. 2013. PMID: 24264462
-
Evolution, immunity and the emergence of brain superautoantigens.F1000Res. 2017 Feb 21;6:171. doi: 10.12688/f1000research.10950.1. eCollection 2017. F1000Res. 2017. PMID: 28529699 Free PMC article.
-
Congenital hypothyroidism mutations affect common folding and trafficking in the α/β-hydrolase fold proteins.FEBS J. 2012 Dec;279(23):4293-305. doi: 10.1111/febs.12019. Epub 2012 Nov 1. FEBS J. 2012. PMID: 23035660 Free PMC article.
References
-
- Di Jeso, B., and Arvan, P. (2004) in The Thyroid (Braverman, L. E., and Utiger, R., eds), 9th Ed., pp. 77–95, Lippincott Williams & Wilkins, Philadelphia, PA
-
- Taurog, A. (1999) Biochimie (Paris) 81 557–562 - PubMed
-
- Marriq, C., Lejeune, P. J., Venot, N., and Vinet, L. (1991) Mol. Cell. Endocrinol. 81 155–164 - PubMed
-
- Dunn, A. D., Corsi, C. M., Myers, H. E., and Dunn, J. T. (1998) J. Biol. Chem. 273 25223–25229 - PubMed
-
- Lamas, L., and Taurog, A. (1977) Endocrinology 100 1129–1136 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
