

An ecological analysis of the herbivory-elicited JA burst and its metabolism: plant memory processes and predictions of the moving target model
- PMID: 19277115
- PMCID: PMC2650097
- DOI: 10.1371/journal.pone.0004697
An ecological analysis of the herbivory-elicited JA burst and its metabolism: plant memory processes and predictions of the moving target model
Abstract
Background: Rapid herbivore-induced jasmonic acid (JA) accumulation is known to mediate many induced defense responses in vascular plants, but little is known about how JA bursts are metabolized and modified in response to repeated elicitations, are propagated throughout elicited leaves, or how they directly influence herbivores.
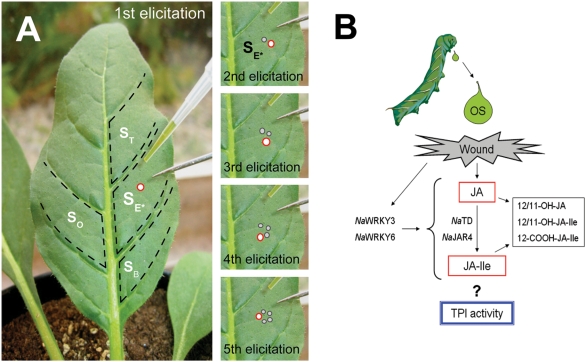
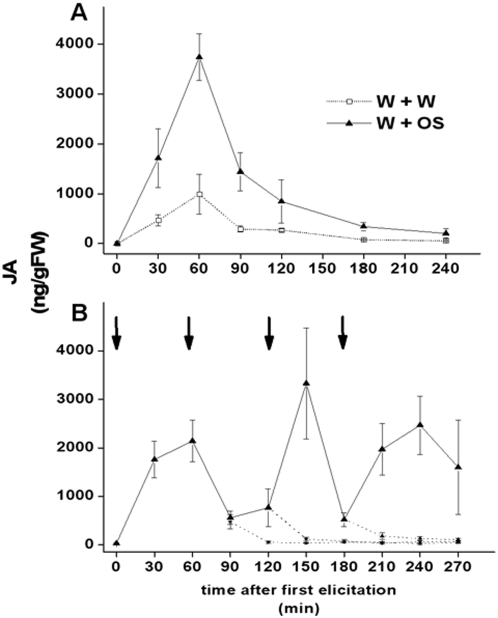
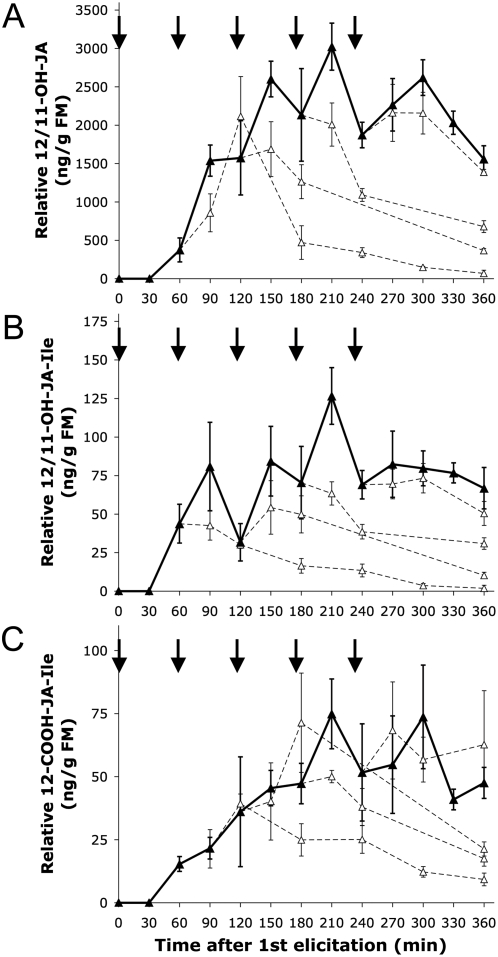
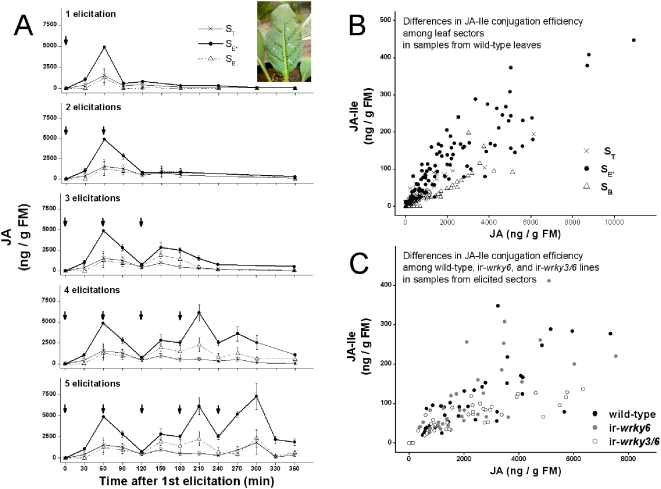
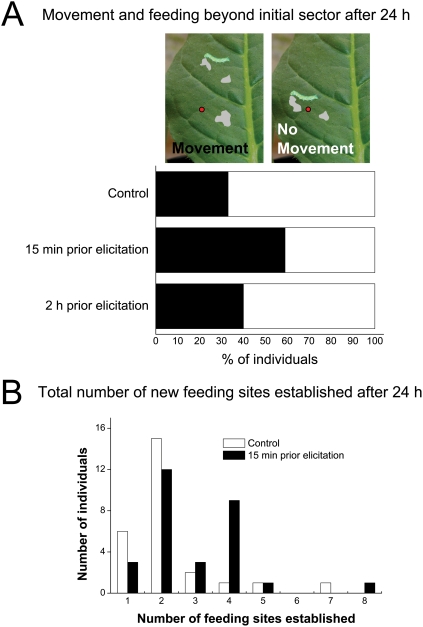
Methodology/principal findings: We found the JA burst in a native population of Nicotiana attenuata to be highly robust despite environmental variation and we examined the JA bursts produced by repeated elicitations with Manduca sexta oral secretions (OS) at whole- and within-leaf spatial scales. Surprisingly, a 2(nd) OS-elicitation suppressed an expected JA burst at both spatial scales, but subsequent elicitations caused more rapid JA accumulation in elicited tissue. The baseline of induced JA/JA-Ile increased with number of elicitations in discrete intervals. Large veins constrained the spatial spread of JA bursts, leading to heterogeneity within elicited leaves. 1(st)-instar M. sexta larvae were repelled by elicitations and changed feeding sites. JA conjugated with isoleucine (JA-Ile) translates elicitations into defense production (e.g., TPIs), but conjugation efficiency varied among sectors and depended on NaWRKY3/6 transcription factors. Elicited TPI activity correlated strongly with the heterogeneity of JA/JA-Ile accumulations after a single elicitation, but not repeated elicitations.
Conclusions/significance: Ecologically informed scaling of leaf elicitation reveals the contribution of repeated herbivory events to the formation of plant memory of herbivory and the causes and importance of heterogeneity in induced defense responses. Leaf vasculature, in addition to transmitting long-distance damage cues, creates heterogeneity in JA bursts within attacked leaves that may be difficult for an attacking herbivore to predict. Such unpredictability is a central tenet of the Moving Target Model of defense, which posits that variability in itself is defensive.
Conflict of interest statement
Figures



References
-
- Reymond P, Farmer EE. Jasmonate and salicylate as global signals for defense gene expression. Current Opinion in Plant Biology. 1998;1:404–411. - PubMed
-
- Rakwal R, Komatsu S. Role of jasmonate in the rice (Oryza sativa L.) self-defense mechanism using proteome analysis. Electrophoresis. 2000;21:2492–2500. - PubMed
-
- Kessler A, Baldwin IT. Plant responses to insect herbivory: the emerging molecular analysis. Annu Rev Plant Biol. 2002;53:299–328. - PubMed
-
- Devoto A, Turner JG. Jasmonate-regulated Arabidopsis stress signalling network. Physiologia Plantarum. 2005;123:161–172.
-
- Zavala JA, Baldwin IT. Jasmonic acid signalling and herbivore resistance traits constrain regrowth after herbivore attack in Nicotiana attenuata. Plant Cell and Environment. 2006;29:1751–1760. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
