

Dynamic dysfunction in dihydrofolate reductase results from antifolate drug binding: modulation of dynamics within a structural state
- PMID: 19278653
- PMCID: PMC2693025
- DOI: 10.1016/j.str.2009.01.005
Dynamic dysfunction in dihydrofolate reductase results from antifolate drug binding: modulation of dynamics within a structural state
Abstract
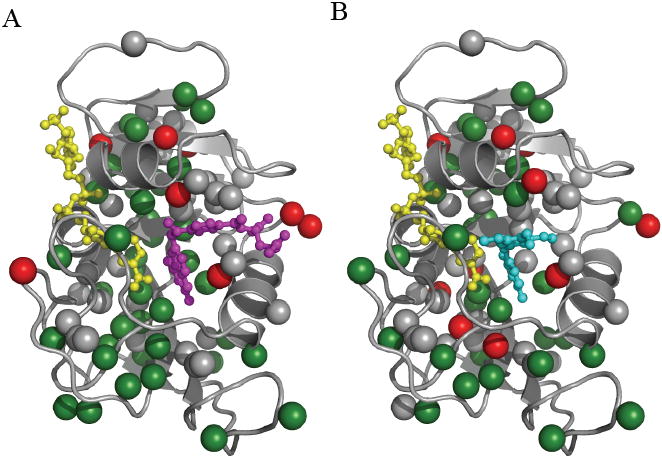
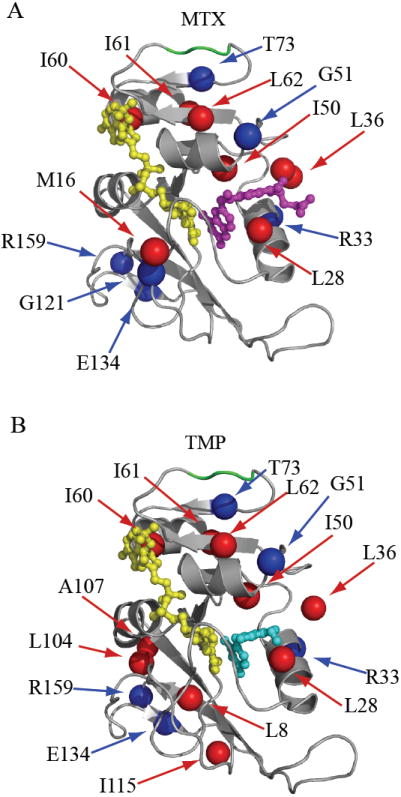
The arduous task of rationally designing small-molecule enzyme inhibitors is complicated by the inherent flexibility of the protein scaffold. To gain insight into the changes in dynamics associated with small-molecule-based inhibition, we have characterized, using NMR spectroscopy, Escherichia coli dihydrofolate reductase in complex with two drugs: methotrexate and trimethoprim. The complexes allowed the intrinsic dynamic effects of drug binding to be revealed within the context of the "closed" structural ensemble. Binding of both drugs results in an identical decoupling of global motion on the micro- to millisecond timescale. Consistent with a change in overall dynamic character, the drugs' perturbations to pico- to nanosecond backbone and side-chain methyl dynamics are also highly similar. These data show that the inhibitors simultaneously modulate slow concerted switching and fast motions at distal regions of dihydrofolate reductase, providing a dynamic link between the substrate binding site and distal loop residues known to affect catalysis.
Figures
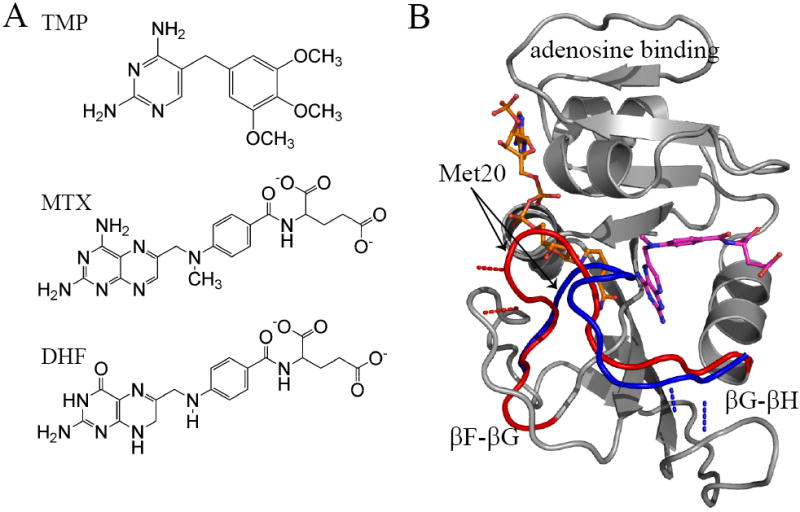
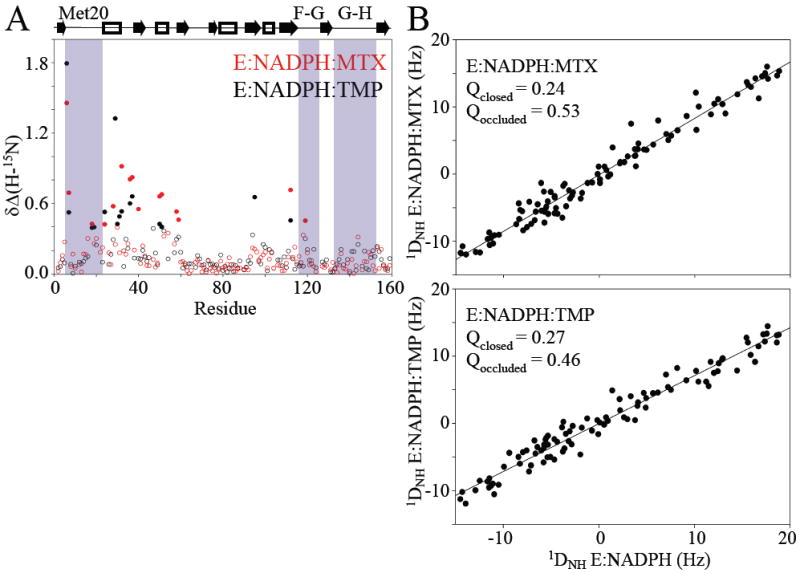
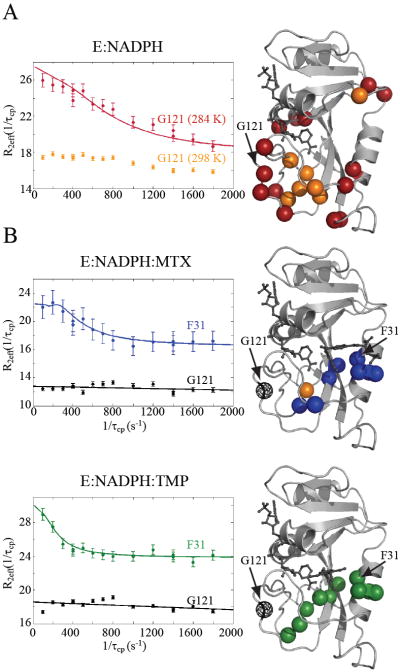
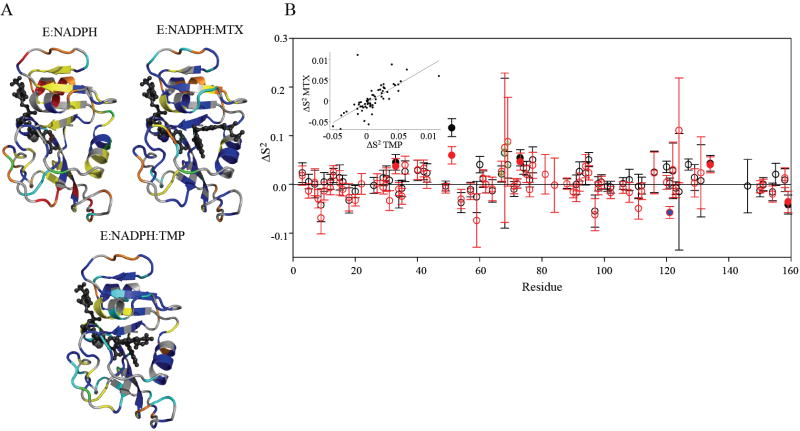
Comment in
-
Communication breakdown: protein dynamics and drug design.Structure. 2009 Mar 11;17(3):319-20. doi: 10.1016/j.str.2009.02.004. Structure. 2009. PMID: 19278644
References
-
- Boehr DD, McElheny D, Dyson HJ, Wright PE. Science. Vol. 313. (New York, N.Y): 2006. The dynamic energy landscape of dihydrofolate reductase catalysis; pp. 1638–1642. - PubMed
-
- Burton LL, Lazo JS, Parker KL. Goodman & Gilman’s the Pharmacological Basis of Therapeutics. 11. New York: McGraw-Hill; 2006.
-
- Cameron CE, Benkovic SJ. Evidence for a functional role of the dynamics of glycine-121 of Escherichia coli dihydrofolate reductase obtained from kinetic analysis of a site-directed mutant. Biochemistry. 1997;36:15792–15800. - PubMed
-
- Careri G, Fasella P, Gratton E. Enzyme dynamics: the statistical physics approach. Annual review of biophysics and bioengineering. 1979;8:69–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
