

Stimulation-induced formation of the reserve pool of vesicles in Drosophila motor boutons
- PMID: 19279147
- PMCID: PMC2681433
- DOI: 10.1152/jn.91122.2008
Stimulation-induced formation of the reserve pool of vesicles in Drosophila motor boutons
Abstract
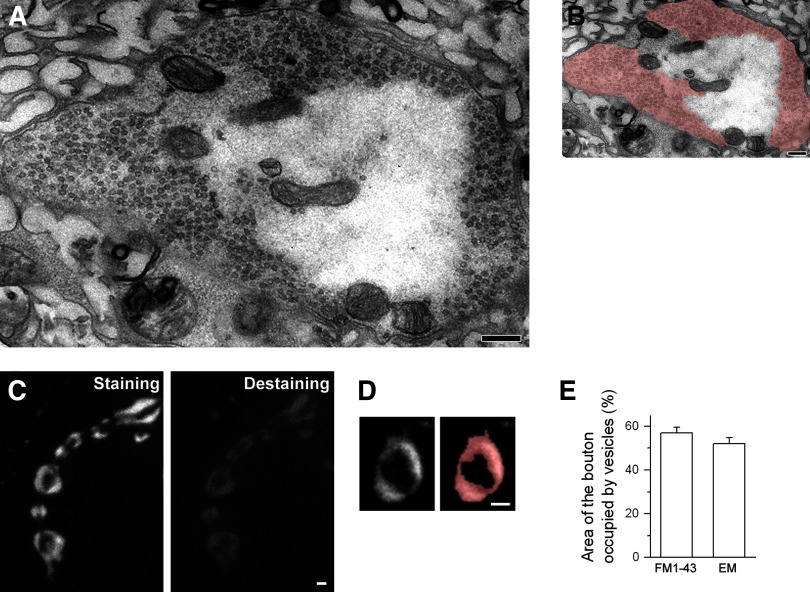
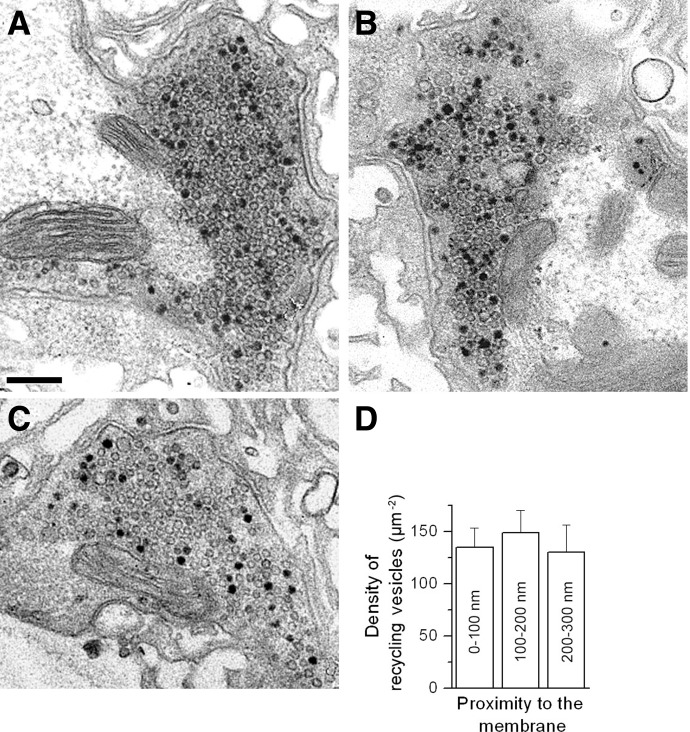
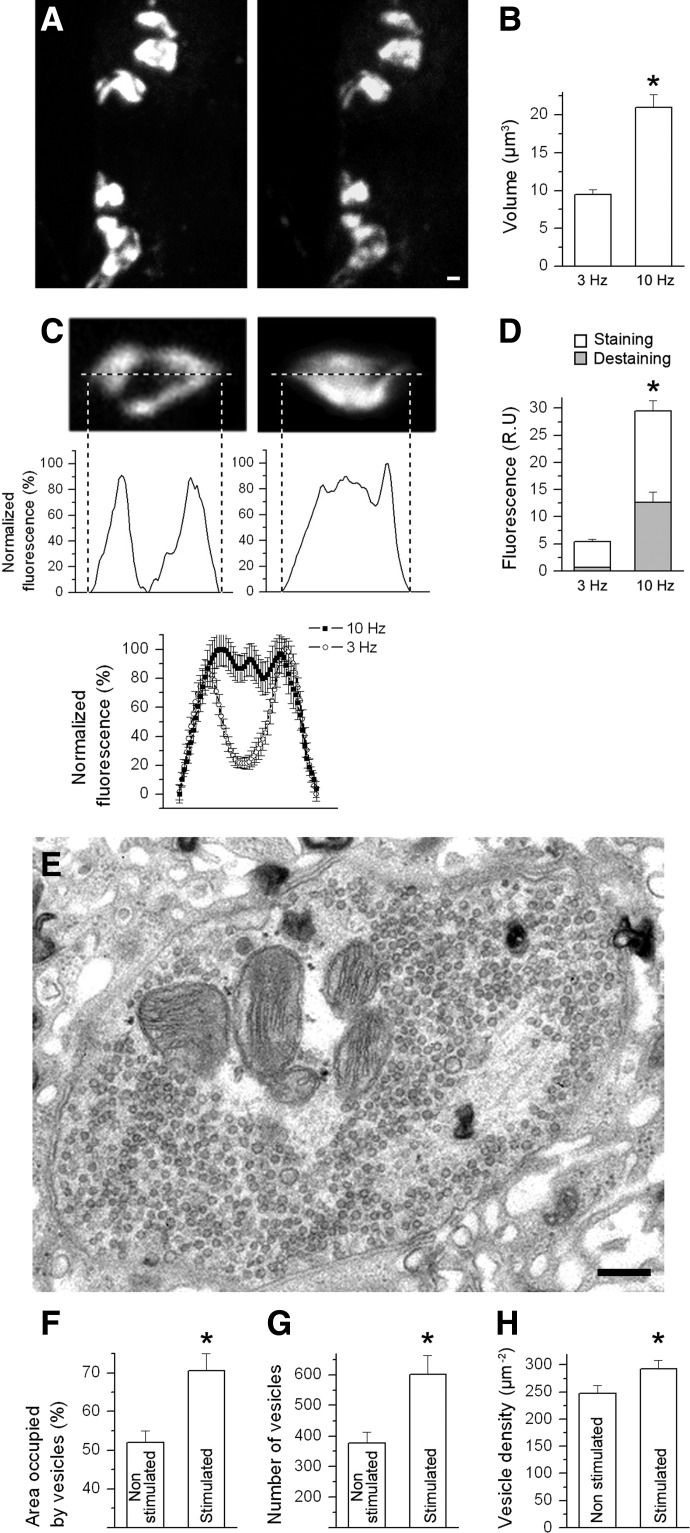
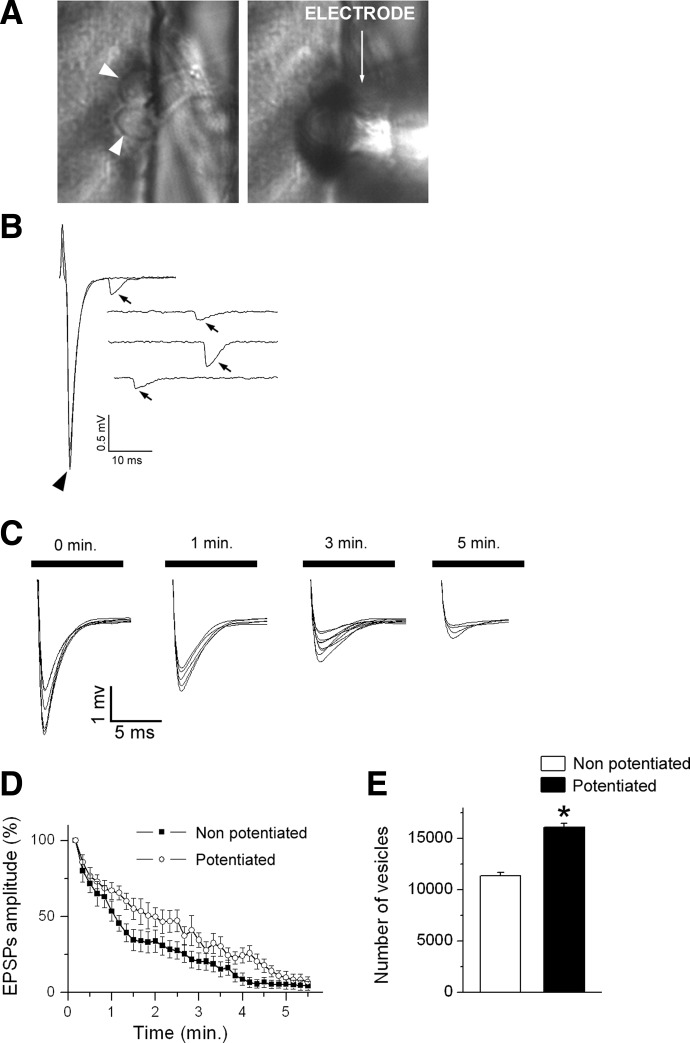
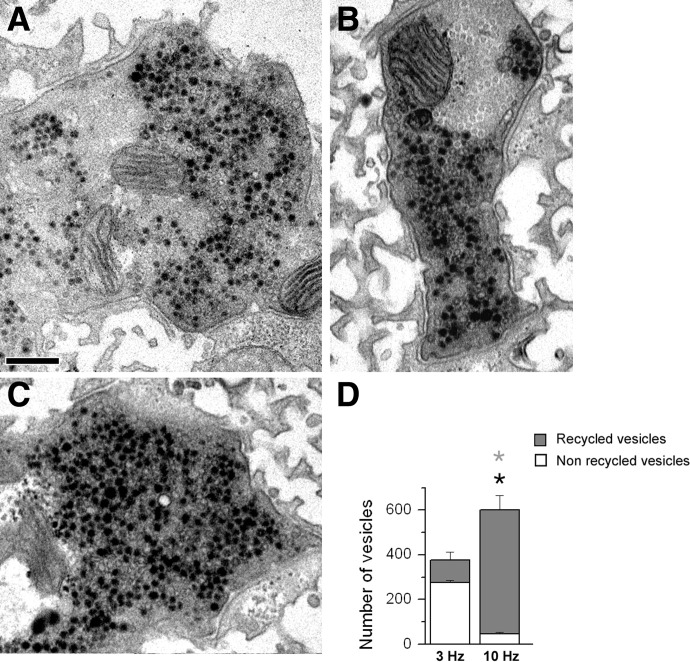
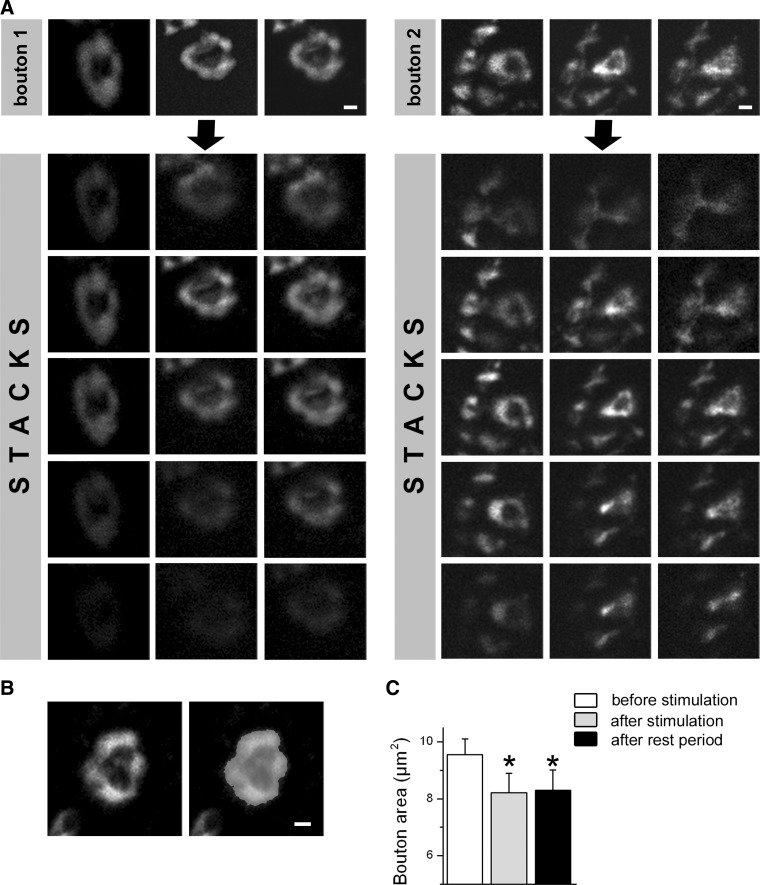
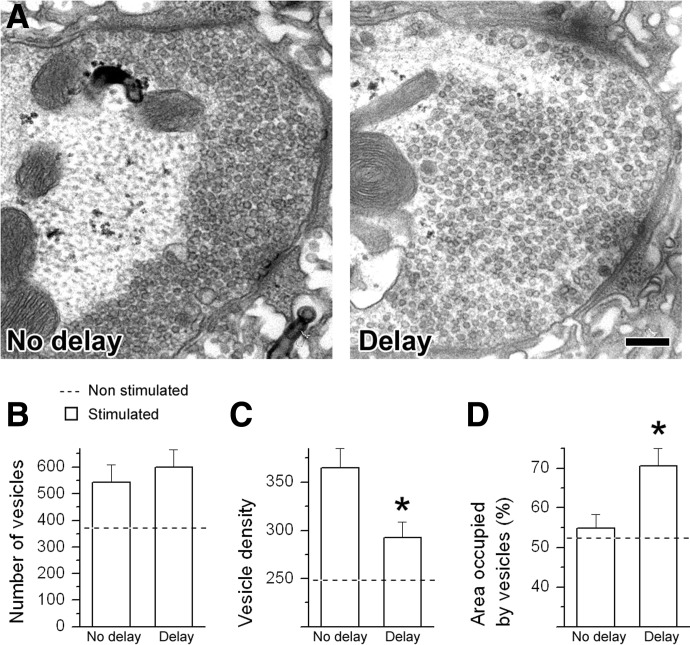
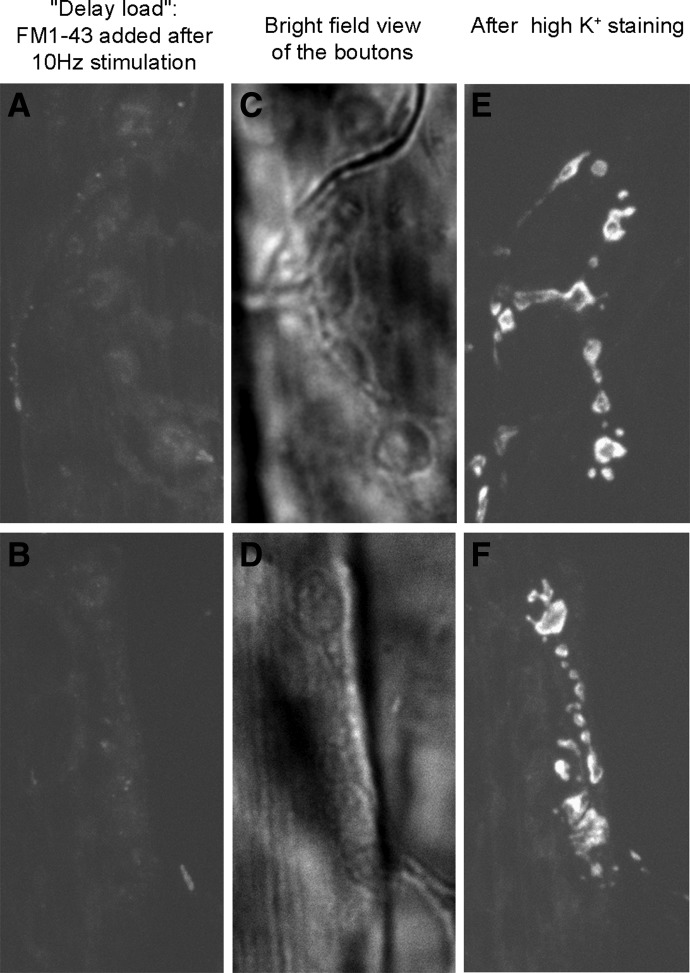
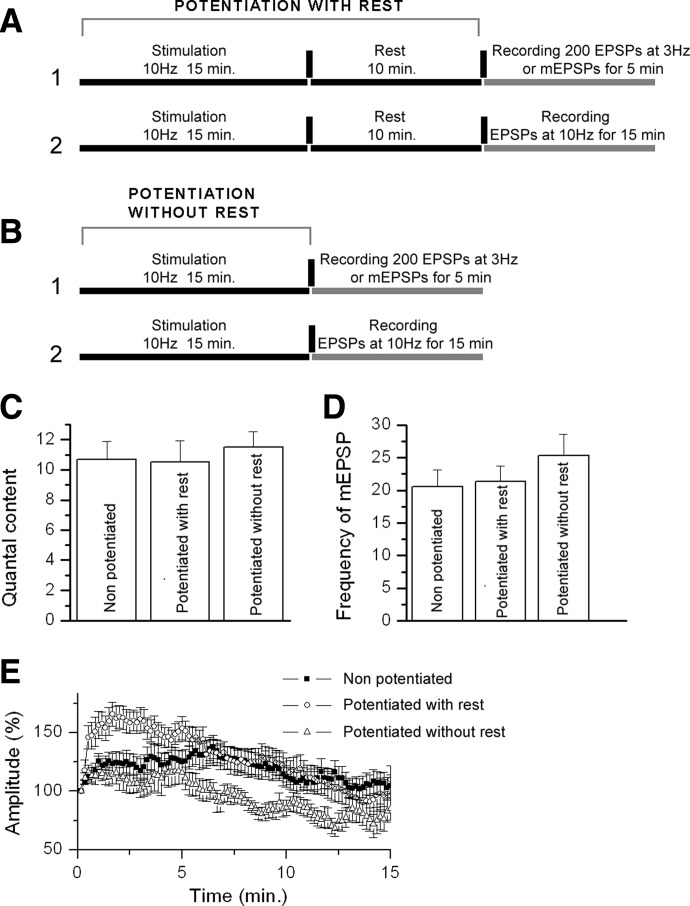
We combined electron microscopy (EM), synaptic vesicle staining by fluorescent marker FM1-43, photoconversion of the dye into an electron dense product, and electrical recordings of synaptic responses to study the distribution of reserve and recycling vesicles and its dependence on stimulation in Drosophila motor boutons. We showed that, at rest, vesicles are distributed over the periphery of the bouton, with the recycling and reserve pools being intermixed and the central core of the bouton being devoid of vesicles. Continuous high-frequency stimulation followed by a resting period mobilized the reserve vesicles into the recycling pool and, most notably, produced an increase in vesicle abundance. Recordings of synaptic activity from the temperature-sensitive endocytosis mutant shibire during continuous stimulation until complete depression provided an independent estimate of the increase in vesicle abundance on intense stimulation. EM analysis demonstrated that continuous stimulation produced an increase in the vesicle density, whereas during a subsequent resting period, vesicles filled empty areas of the bouton, spreading toward its central core. Although the observed structural potentiation did not alter basal transmitter release, it produced an increased synaptic enhancement during high-frequency stimulation. The latter effect was not observed when the boutons were potentiated using high-frequency stimulation without a subsequent resting period. We concluded therefore that the newly formed vesicles replenish the reserve pool during a resting period following intense stimulation.
Figures
References
-
- Akbergenova Y, Bykhovskaia M. Synapsin maintains the reserve vesicle pool and spatial segregation of the recycling pool in Drosophila presynaptic boutons. Brain Res 1178: 52–64, 2007. - PubMed
-
- Atwood HL, Govind CK, Wu CF. Differential ultrastructure of synaptic terminals on ventral longitudinal abdominal muscles in Drosophila larvae. J Neurobiol 24: 1008–1024, 1993. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases