

doi: 10.4161/org.4.2.5858.
Wnt signaling in breast organogenesis
Affiliations
- PMID: 19279723
- PMCID: PMC2634257
- DOI: 10.4161/org.4.2.5858
Item in Clipboard
Wnt signaling in breast organogenesis
Organogenesis.
2008 Apr.
Abstract
Wnt signals play a critical role in regulating the normal development of the mammary gland and dysregulation of Wnt signaling causes breast cancer. This pathway is involved in the earliest development of the mammary gland in embryos and its role extends through the functional differentiation of the gland during pregnancy. In this review, we summarize the molecular mechanisms through which Wnts regulate mammary gland development in the mouse.
Keywords: Wnt; cancer; embryo; mammary gland; postnatal; stem cell.
Figures
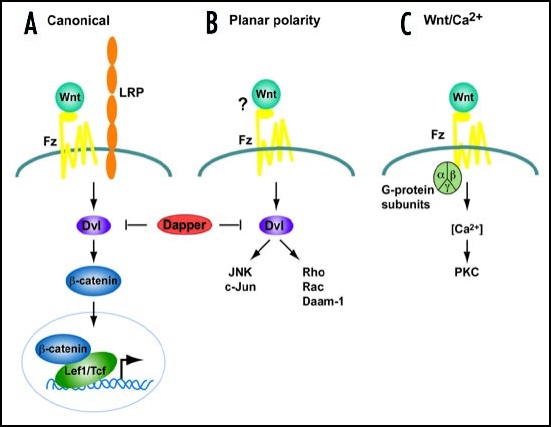
Simplified schemes of the three Wnt signaling cascades.
(A) Canonical Pathway. A Wnt ligand binds to a Frizzled receptor and a co-receptor of low density lipoprotein (Lrp) family. Dishevelled (Dvl) and β-catenin are required to transduce the Wnt signal, leading to a transcriptional response mediated by transcription factors of the Tcf/Lef1 family. (B) Planar cell Polarity pathway. In this pathway, Dvl also transduces signals although independent of β-catenin activating JNK and Rho kinase cascades. (C) The Wnt/Ca2+ pathway signals via heterotrimeric G-proteins to mobilize intracellular Ca2+ and stimulate protein kinase C (PKC). The requirement for Dvl is controversial. In vertebrates this pathway is activated by the same ligands as (B). Dapper, a Dvl-associated antogonist, antagonizes both (A and B) pathways.
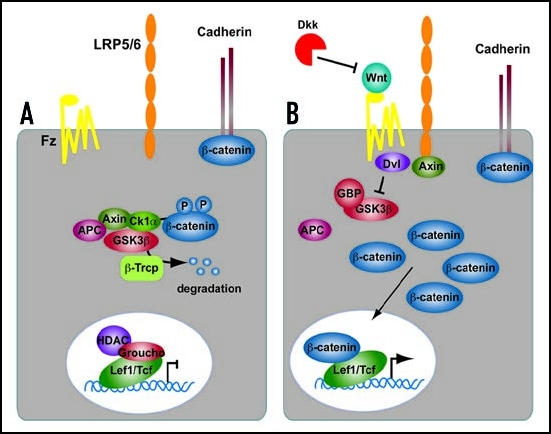
Summary of the Canonical Wnt signaling cascade. (A) In the absence of a Wnt ligand, β-catenin is degraded through interactions between APC, Axin and the protein kinase GSK3β. In the nucleus, Lef1/Tcf transcription factors are in complexes with corepressors (e.g., CtBP, HDAC, Groucho) thereby repressing the transcription of Wnt target genes. (B) Wnt proteins bind to the Frizzled/Lrp receptor complex at the cell surface. These receptors transduce a signal to Dishevelled (Dvl) and to Axin, which may directly interact. As a consequence, β-catenin degradation is inhibited, and it accumulates in the cytoplasm and nucleus. β-catenin interacts with Lef1/Tcfs in the nucleus to control transcription of Wnt target genes. The secreted antagonist Dickkopf (Dkk) does not bind to Wnt ligands directly but prevents formation of an active Wnt-Fzd-Lrp complex.
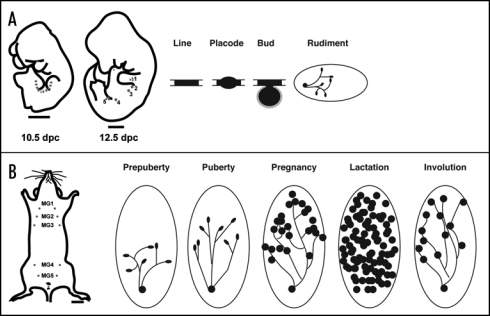
Morphological and schematic development of the mouse mammary gland during embryogenesis and postnatal life. (A) Cartoon illustrating the mammary line at 10.5 dpc in the mouse embryo, and the position of mammary rudiments in the 12.5 dpc mouse embryo. The mammary line forms transiently along the anterior-posterior axis at each flank of the embryo, eventually giving rise to 5 pairs of mammary placodes. There are three thoracic (1–3) and two inguinal (4 and 5) mammary buds. These buds give rise to a rudimental ductal tree by birth. (B) In the adult female mouse five pairs of mammary glands are found: three thoracic gland (1–3) and two inguinal glands (4 and 5). During puberty, hormones induce the rudimentry ductal network to elongate and proliferate. The ends of growing ducts form terminal end buds (TEB) during puberty (solid black ovals). The large oval shown in the postnatal stages depicts the mammary fat pad (stroma). During pregnancy, alveolar proliferation and differentiation occurs to produce milk proteins for mammary function during lactation. Mammary ducts are shown as solid lines and the lobuloalveolar structures are presented as solid black circles. Tissue remodeling and cell death occur during involution and bring the gland back to a virgin-like resting state. Scale bars in (A) embryo 1000 mm; (B) adult, 2500 mm.
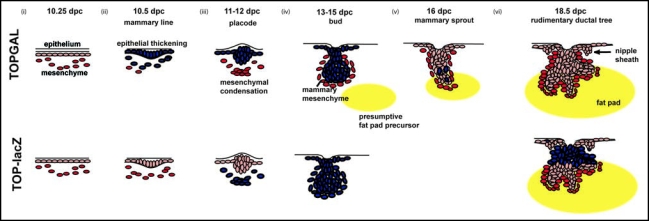
Schematic overview of Wnt signaling during embryonic mammary gland development in the mouse. Initiation of mammary gland development proceeds through three inductive epithelial-mesenchymal signals. (i) Embryonic dermis instructs the overlying ectoderm to initiate placode formation.
(ii) Epithelial placode signals to mesenchymal fibroblasts to form mesenchymal condensations. (iii) Mesenchymal condensations signal to the overlying bud to stimulate proliferation and downgrowth into the developing dermis. (iv) Subsequently, as the bud invaginates into the underlying mesenchyme, mammary mesenchymal cells surround the developing bud. A secondary mesenchyme, which gives rise to the future fat pad begins to develop. (v) The mammary sprout continues to proliferate and grow downward into the presumptive fat pad giving rise to a rudimentary ductal structure of 15–20 branches (vi). The nipple sheath also develops from the epidermal cells. Blue cells represent Wnt signalling activity during mammary gland development in TOPGAL (top) and TOP-lacZ transgenic embryos.
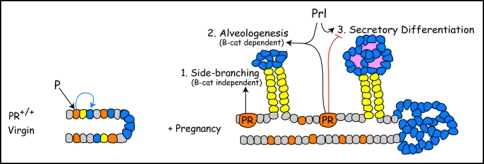
Proposed interactions between progesterone receptor (PR) and β-catenin signaling during alveologenesis. In the normal virgin mammary gland, PR induces a non-uniform PR expression pattern and competence to respond to β-catenin in alveolar progenitors (blue) during ductal maturation. During early pregnancy, PR-Wnt4 signaling induces expansion of ductal progenitors (yellow) to form side-branches through βcatenin-independent routes. PR and PRL (prolactin) cooperate to induce alveologenesis, a process that is β-catenin-dependent. Later in pregnancy, PR restrains and PRL promote alveolar differentiation. β-catenin is required for alveologenesis and may participate at multiple steps in the secretory differentiation pathway.
References
-
- Moon RT, Shah K. Developmental biology: Signaling polarity. Nature. 2002;417:239–240. - PubMed
-
- Bienz M, Clevers H. Armadillo/beta-catenin signals in the nucleus-proof beyond a reasonable doubt? Nat Cell Biol. 2003;5:179–182. - PubMed
-
- Alonso L, Fuchs E. Stem cells in the skin: waste not, Wnt not. Genes Dev. 2003;17:1189–1200. - PubMed
-
- Staal FJ, Clevers HC. Wnt signaling in the thymus. Curr Opin Immunol. 2003;15:204–208. - PubMed
-
- Brennan KR, Brown AM. Wnt proteins in mammary development and cancer. J Mammary Gland Biol Neoplasia. 2004;9:119–131. - PubMed
LinkOut - more resources
Full Text Sources