

Hybrid coarse-graining approach for lipid bilayers at large length and time scales
- PMID: 19281167
- PMCID: PMC2689158
- DOI: 10.1021/jp8087868
Hybrid coarse-graining approach for lipid bilayers at large length and time scales
Abstract
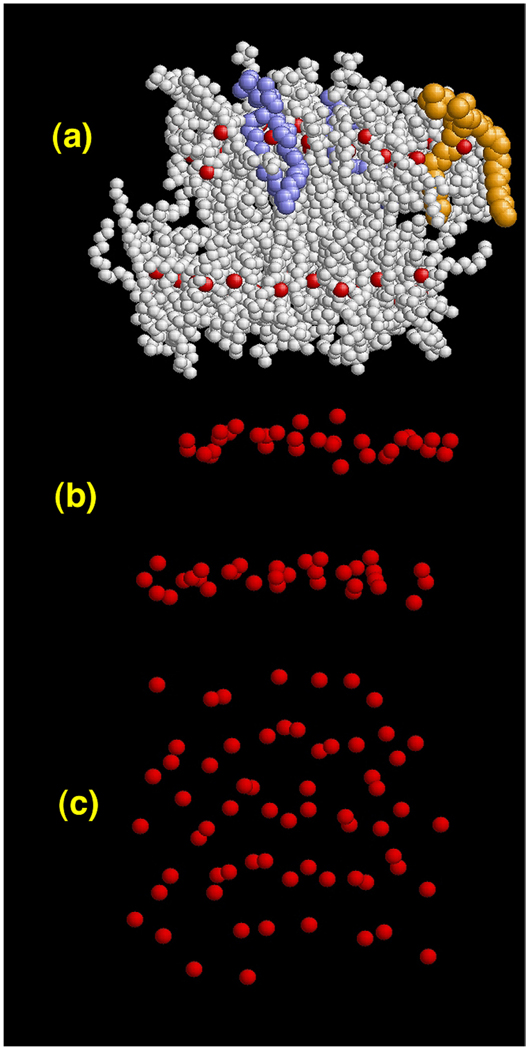
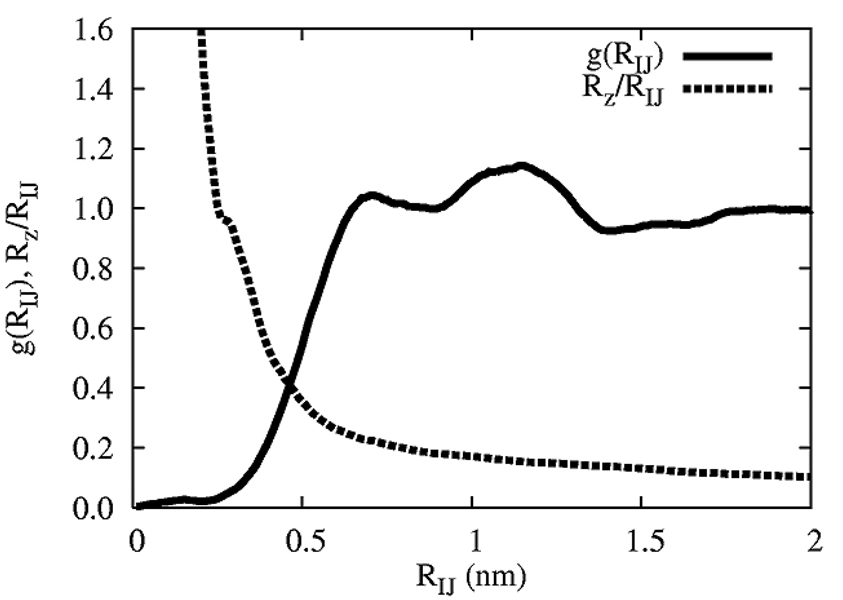
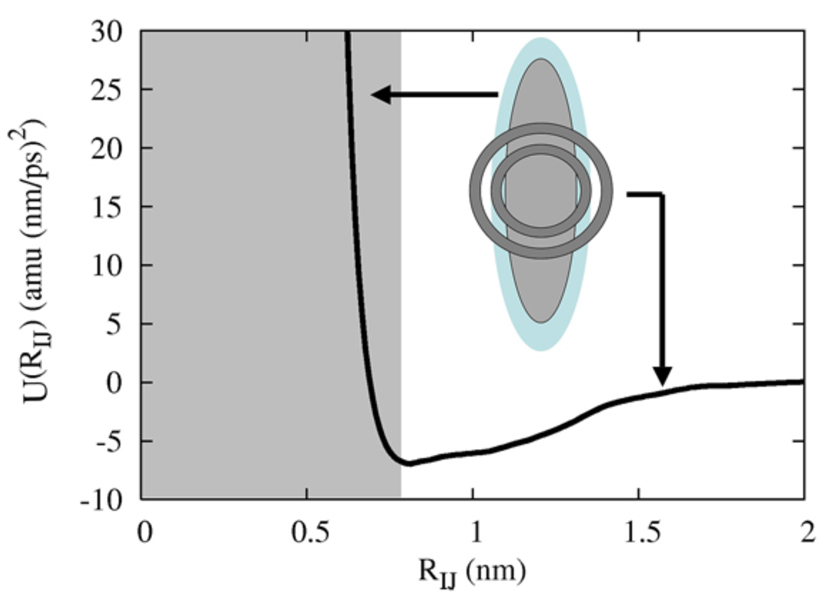
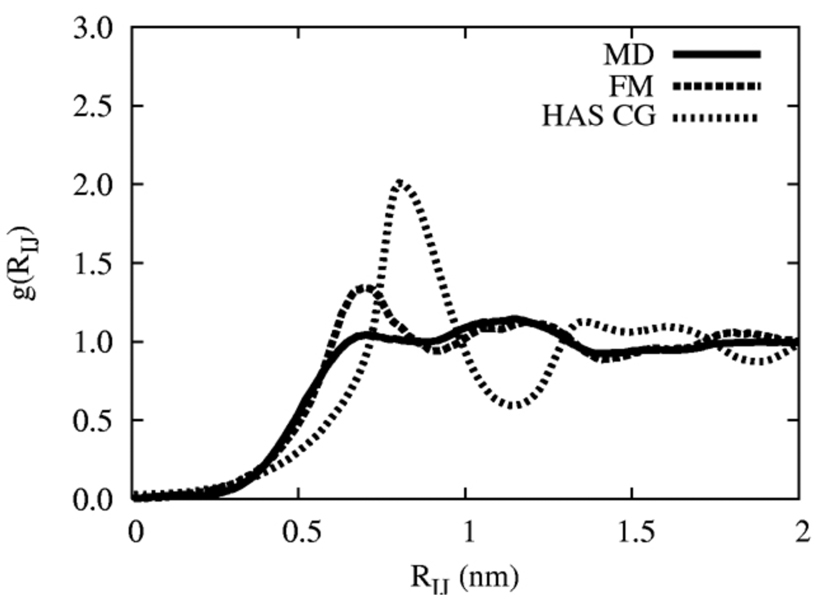
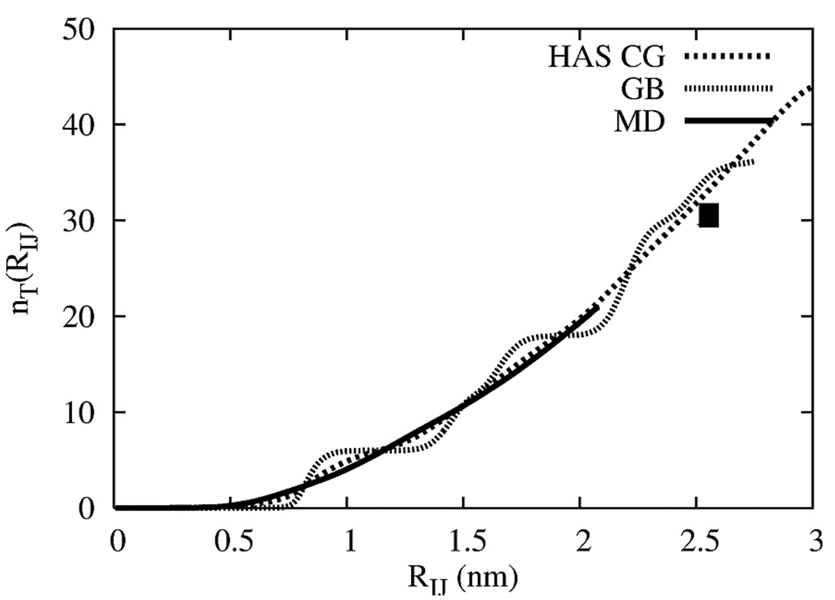
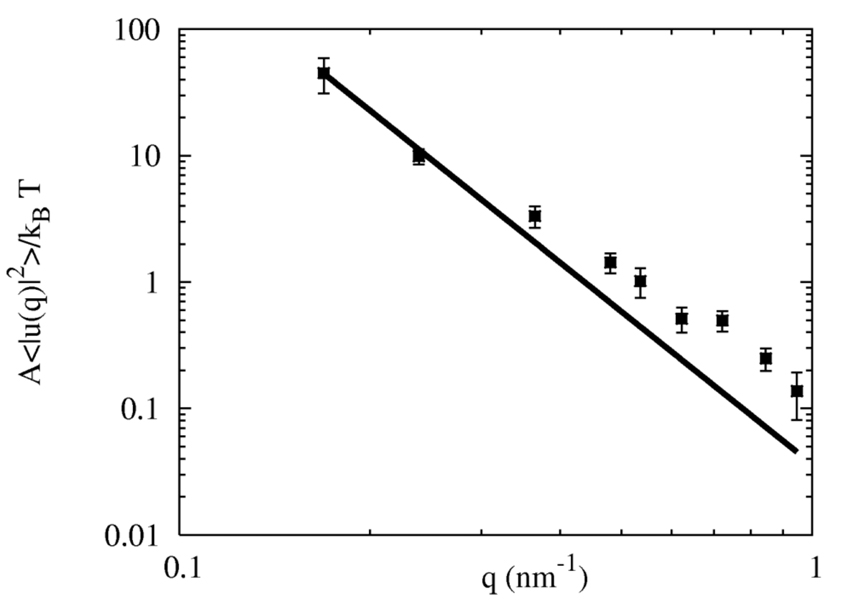
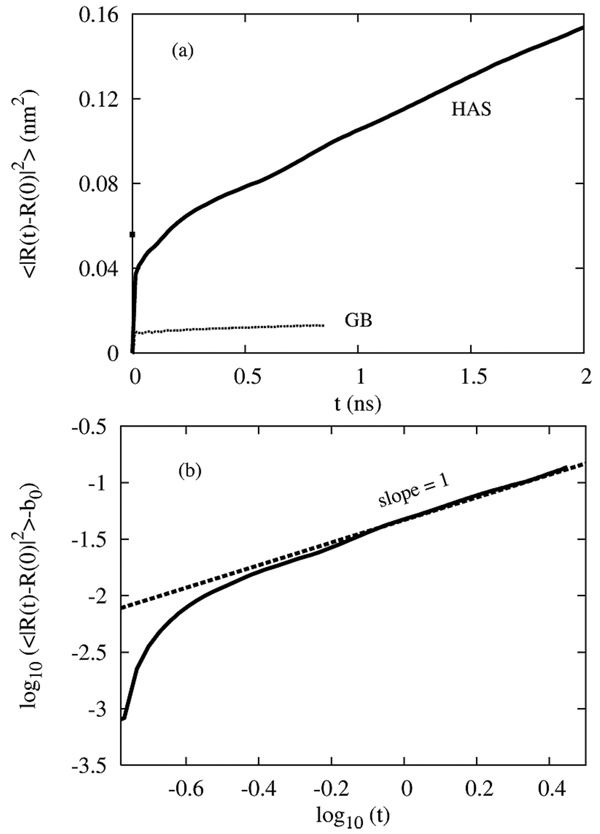
A hybrid analytic-systematic (HAS) coarse-grained (CG) lipid model is developed and employed in a large-scale simulation of a liposome. The methodology is termed hybrid analytic-systematic because one component of the interaction between CG sites is variationally determined from the multiscale coarse-graining (MS-CG) methodology, whereas the remaining component utilizes an analytic potential. The systematic component models the in-plane center-of-mass interaction of the lipids as determined from an atomistic-level MD simulation of a bilayer. The analytic component is based on the well-known Gay-Berne ellipsoid-of-revolution liquid-crystal model and is designed to model the highly anisotropic interactions at a highly coarse-grained level. The HAS CG approach is the first step in an "aggressive" CG methodology designed to model multicomponent biological membranes at very large length and time scales.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
