

Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor
- PMID: 19285469
- PMCID: PMC2671566
- DOI: 10.1016/j.neuron.2008.12.025
Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor
Abstract
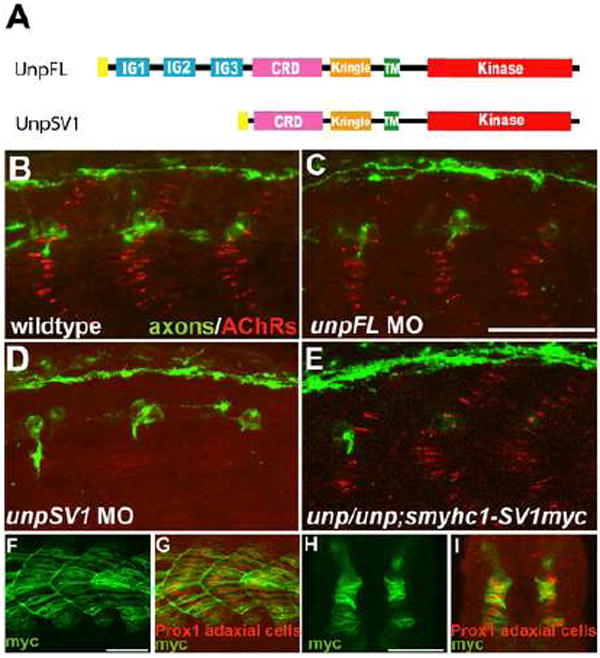
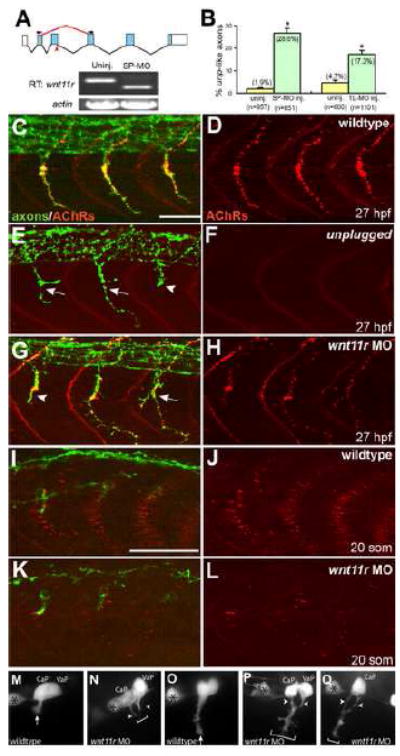
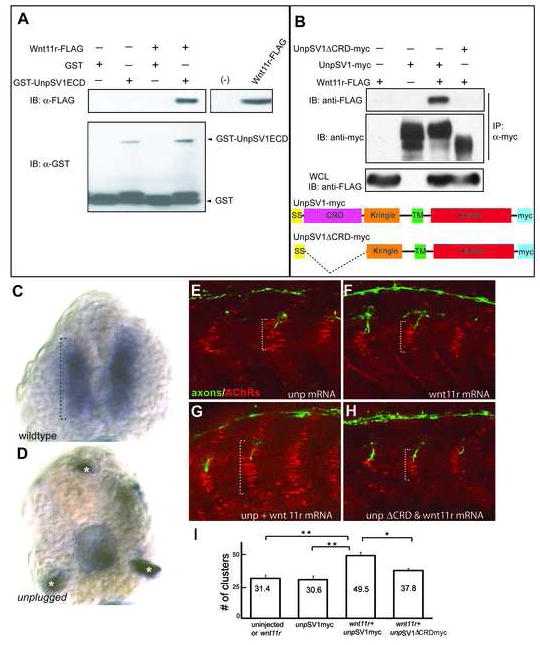
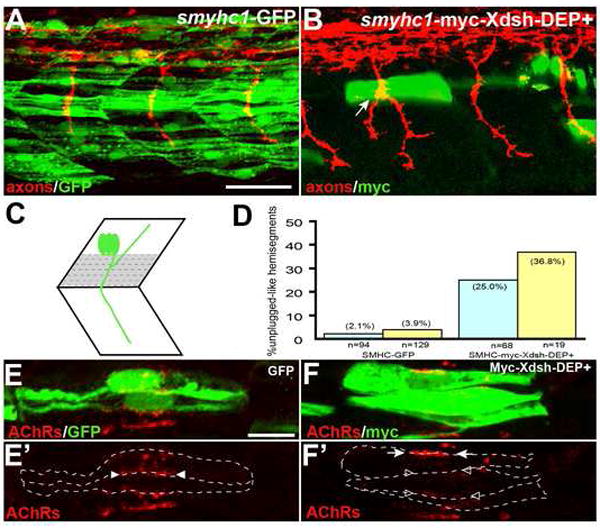
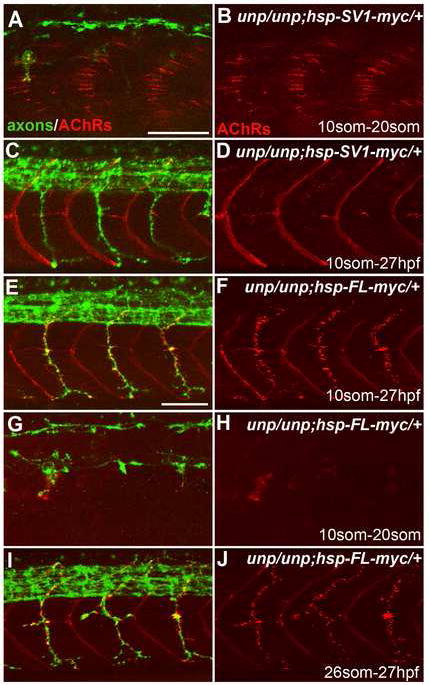
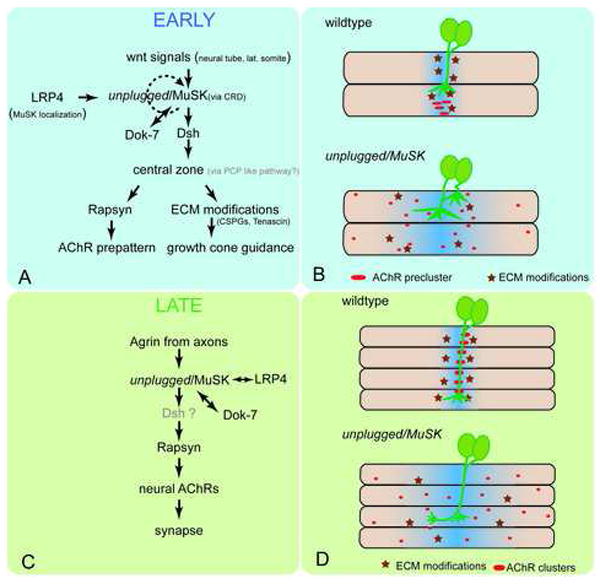
Early during neuromuscular development, acetylcholine receptors (AChRs) accumulate at the center of muscle fibers, precisely where motor growth cones navigate and synapses eventually form. Here, we show that Wnt11r binds to the zebrafish unplugged/MuSK ectodomain to organize this central muscle zone. In the absence of such a zone, prepatterned AChRs fail to aggregate and, as visualized by live-cell imaging, growth cones stray from their central path. Using inducible unplugged/MuSK transgenes, we show that organization of the central muscle zone is dispensable for the formation of neural synapses, but essential for AChR prepattern and motor growth cone guidance. Finally, we show that blocking noncanonical dishevelled signaling in muscle fibers disrupts AChR prepatterning and growth cone guidance. We propose that Wnt ligands activate unplugged/MuSK signaling in muscle fibers to restrict growth cone guidance and AChR prepatterns to the muscle center through a mechanism reminiscent of the planar cell polarity pathway.
Figures

References
-
- Burden SJ. Wnts as retrograde signals for axon and growth cone differentiation. Cell. 2000;100:495–497. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
