

Cytochrome c/cardiolipin relations in mitochondria: a kiss of death
- PMID: 19285551
- PMCID: PMC2732771
- DOI: 10.1016/j.freeradbiomed.2009.03.004
Cytochrome c/cardiolipin relations in mitochondria: a kiss of death
Abstract
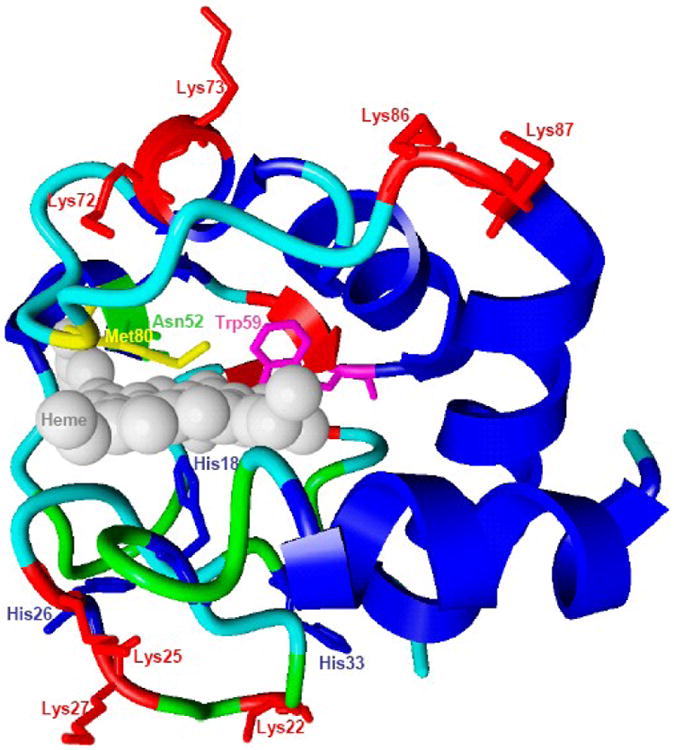
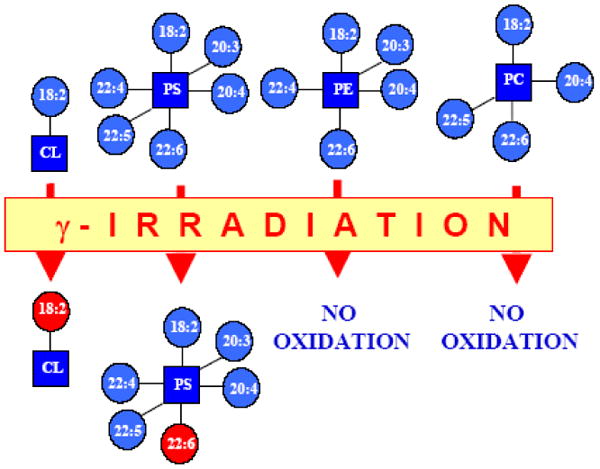
Recently, phospholipid peroxidation products gained a reputation as key regulatory molecules and participants in oxidative signaling pathways. During apoptosis, a mitochondria-specific phospholipid, cardiolipin (CL), interacts with cytochrome c (cyt c) to form a peroxidase complex that catalyzes CL oxidation; this process plays a pivotal role in the mitochondrial stage of the execution of the cell death program. This review is focused on redox mechanisms and essential structural features of cyt c's conversion into a CL-specific peroxidase that represent an interesting and maybe still unique example of a functionally significant ligand change in hemoproteins. Furthermore, specific characteristics of CL in mitochondria--its asymmetric transmembrane distribution and mechanisms of collapse, the regulation of its synthesis, remodeling, and fatty acid composition--are given significant consideration. Finally, new concepts in drug discovery based on the design of mitochondria-targeted inhibitors of cyt c/CL peroxidase and CL peroxidation with antiapoptotic effects are presented.
Figures




References
-
- Halliwell B, Gutteridge J. Free Radicals in Biology and Medicine. Oxford, U.K.: Clarendon Press; 1999.
-
- Skulachev VP. Cytochrome c in the apoptotic and antioxidant cascades. FEBS Lett. 1998;423:275–280. - PubMed
-
- Liu X, Kim CN, Yang J, Jemmerson R, Wang X. Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell. 1996;86:147–157. - PubMed
-
- Passardi F, Theiler G, Zamocky M, Cosio C, Rouhier N, Teixera F, Margis-Pinheiro M, Ioannidis V, Penel C, Falquet L, Dunand C. PeroxiBase: the peroxidase database. Phytochemistry. 2007;68:1605–11. - PubMed
-
- Rouzer CA, Marnett LJ. Mechanism of free radical oxygenation of polyunsaturated fatty acids by cyclooxygenases. Chem Rev. 2003;103:2239–304. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
