

Resampling and editing of mischarged tRNA prior to translation elongation
- PMID: 19285947
- PMCID: PMC2944653
- DOI: 10.1016/j.molcel.2009.01.031
Resampling and editing of mischarged tRNA prior to translation elongation
Abstract
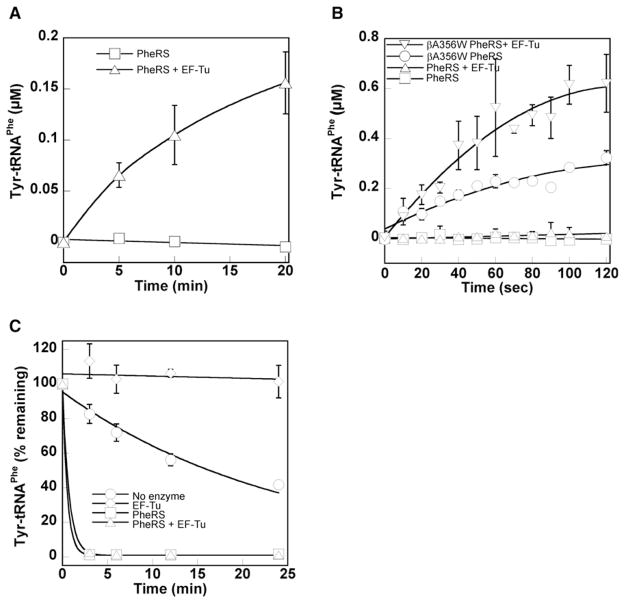
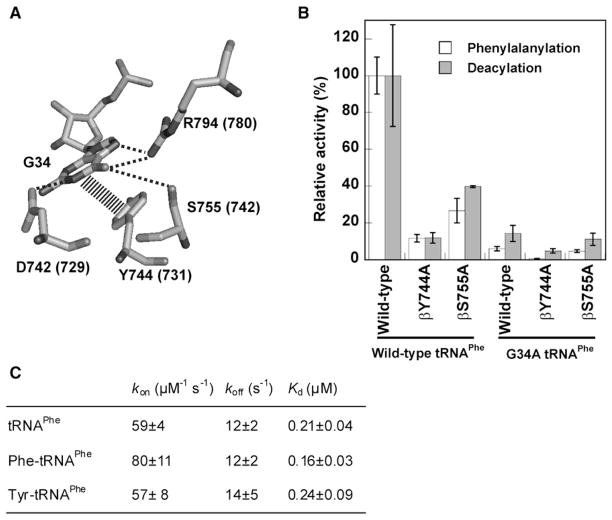
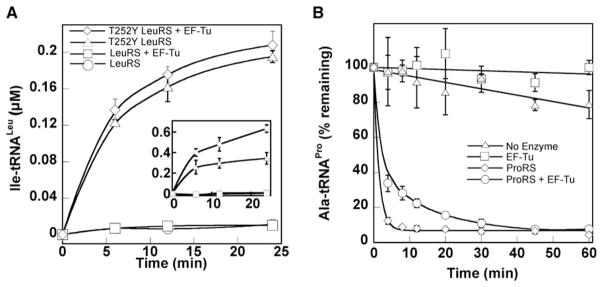
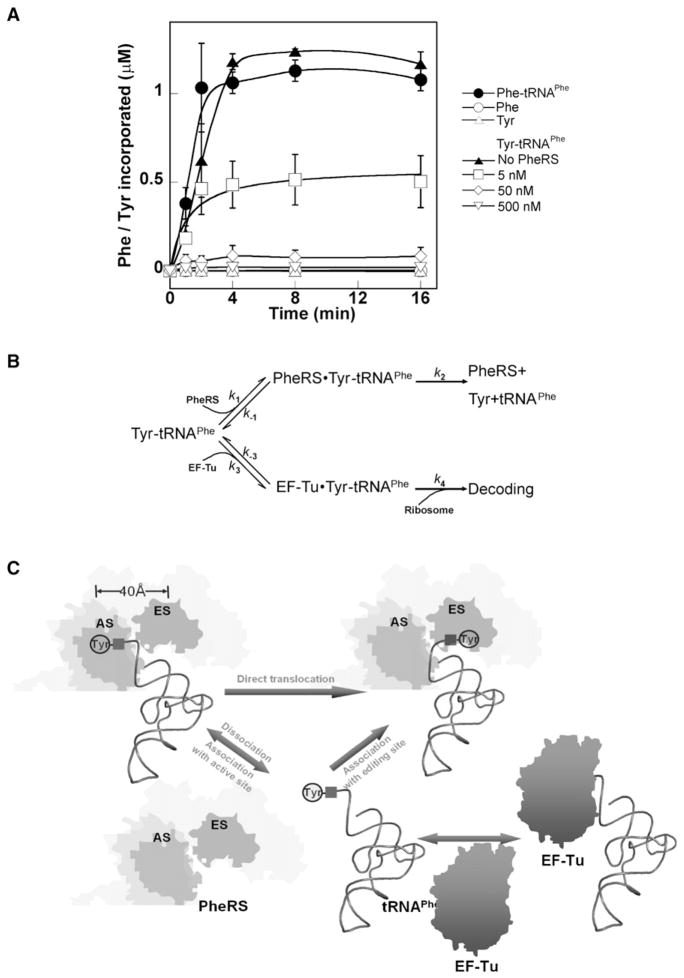
Faithful translation of the genetic code depends on the GTPase EF-Tu delivering correctly charged aminoacyl-tRNAs to the ribosome for pairing with cognate codons. The accurate coupling of cognate amino acids and tRNAs by the aminoacyl-tRNA synthetases is achieved through a combination of substrate specificity and product editing. Once released by aminoacyl-tRNA synthetases, both cognate and near-cognate aminoacyl-tRNAs were considered to be committed to ribosomal protein synthesis through their association with EF-Tu. Here we show instead that aminoacyl-tRNAs in ternary complex with EF-Tu*GTP can readily dissociate and rebind to aminoacyl-tRNA synthetases. For mischarged species, this allows resampling by the product editing pathway, leading to a reduction in the overall error rate of aminoacyl-tRNA synthesis. Resampling of mischarged tRNAs was shown to increase the accuracy of translation over ten fold during in vitro protein synthesis, supporting the presence of an additional quality control step prior to translation elongation.
Figures
References
-
- An S, Musier-Forsyth K. Trans-editing of Cys-tRNAPro by Haemophilus influenzae YbaK protein. J Biol Chem. 2004;279:42359–42362. - PubMed
-
- An S, Musier-Forsyth K. Cys-tRNAPro editing by Haemophilus influenzae YbaK via a novel synthetase. YbaK.tRNA ternary complex. J Biol Chem. 2005;280:34465–34472. - PubMed
-
- Baldwin AN, Berg P. Transfer ribonucleic acid-induced hydrolysis of valyladenylate bound to isoleucyl ribonucleic acid synthetase. J Biol Chem. 1966;241:839–845. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
