

Intracellular sterol dynamics
- PMID: 19286471
- PMCID: PMC2696574
- DOI: 10.1016/j.bbalip.2009.03.002
Intracellular sterol dynamics
Abstract
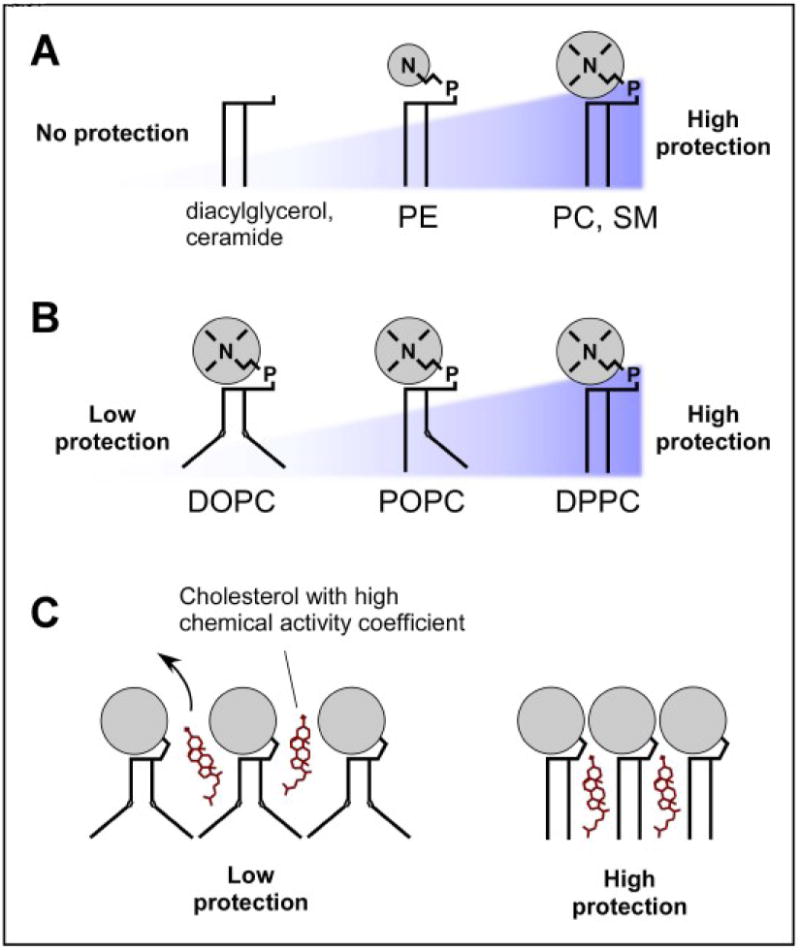
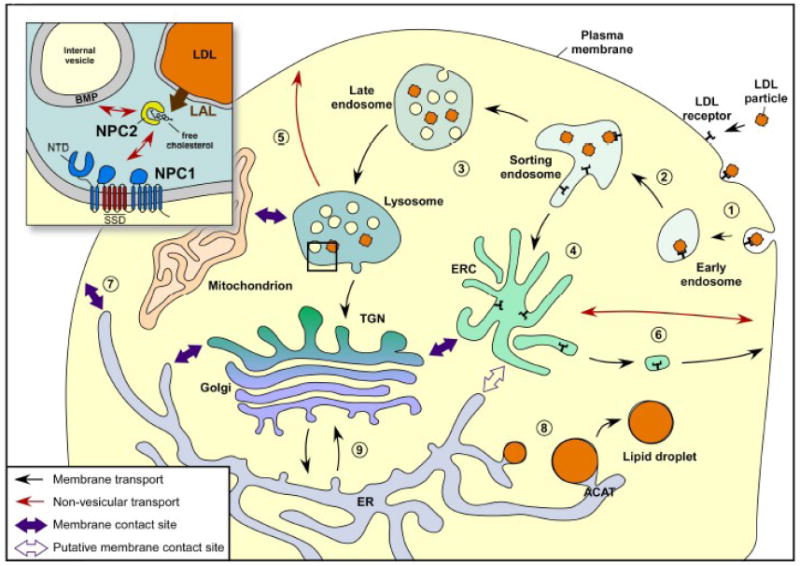
We review the cellular mechanisms implicated in cholesterol trafficking and distribution. Recent studies have provided new information about the distribution of sterols within cells, including analysis of its transbilayer distribution. The cholesterol interaction with other lipids and its engagement in various trafficking processes will determine its proper level in a specific membrane; making the cholesterol distribution uneven among the various intracellular organelles. The cholesterol content is important since cholesterol plays an essential role in membranes by controlling their physicochemical properties as well as key cellular events such as signal transduction and protein trafficking. Cholesterol movement between cellular organelles is highly dynamic, and can be achieved by vesicular and non-vesicular processes. Various studies have analyzed the proteins that play a significant role in these processes, giving us new information about the relative importance of these two trafficking pathways in cholesterol transport. Although still poorly characterized in many trafficking routes, several potential sterol transport proteins have been described in detail; as a result, molecular mechanisms for sterol transport among membranes start to be appreciated.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
