

Regulation of nitrite transport in red blood cells by hemoglobin oxygen fractional saturation
- PMID: 19286940
- PMCID: PMC2685350
- DOI: 10.1152/ajpheart.01303.2008
Regulation of nitrite transport in red blood cells by hemoglobin oxygen fractional saturation
Abstract
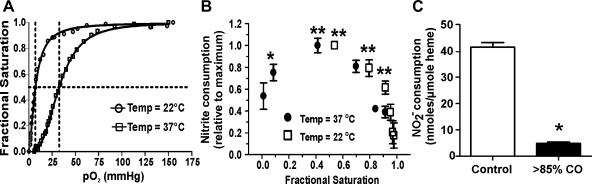
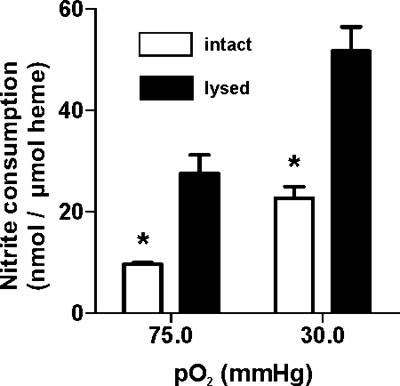
Allosteric regulation of nitrite reduction by deoxyhemoglobin has been proposed to mediate nitric oxide (NO) formation during hypoxia. Nitrite is predominantly an anion at physiological pH, raising questions about the mechanism by which it enters the red blood cell (RBC) and whether this is regulated and coupled to deoxyhemoglobin-mediated reduction. We tested the hypothesis that nitrite transport by RBCs is regulated by fractional saturation. Using human RBCs, nitrite consumption was faster at lower fractional saturations, consistent with faster reactions with deoxyheme. A membrane-based regulation was suggested by slower nitrite consumption with intact versus lysed RBCs. Interestingly, upon nitrite addition, intracellular nitrite concentrations attained a steady state that, despite increased rates of consumption, did not change with decreasing oxygen tensions, suggesting a deoxygenation-sensitive step that either increases nitrite import or decreases the rate of nitrite export. A role for anion exchanger (AE)-1 in the control of nitrite export was suggested by increased intracellular nitrite concentrations in RBCs treated with DIDS. Moreover, deoxygenation decreased steady-state levels of intracellular nitrite in AE-1-inhibited RBCs. Based on these data, we propose a model in which deoxyhemoglobin binding to AE-1 inhibits nitrite export under low oxygen tensions allowing for the coupling between deoxygenation and nitrite reduction to NO along the arterial-to-venous gradient.
Figures





References
-
- Adragna NC, Lauf PK. Role of nitrite, a nitric oxide derivative, in K-Cl cotransport activation of low-potassium sheep red blood cells. J Membr Biol 166: 157–167, 1998. - PubMed
-
- Barvitenko NN, Adragna NC, Weber RE. Erythrocyte signal transduction pathways, their oxygenation dependence and functional significance. Cell Physiol Biochem 15: 1–18, 2005. - PubMed
-
- Basu S, Grubina R, Huang J, Conradie J, Huang Z, Jeffers A, Jiang A, He X, Azarov I, Seibert R, Mehta A, Patel R, King SB, Hogg N, Ghosh A, Gladwin MT, Kim-Shapiro DB. Catalytic generation of N2O3 by the concerted nitrite reductase and anhydrase activity of hemoglobin. Nat Chem Biol 3: 785–794, 2007. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous