

p53-cofactor JMY is a multifunctional actin nucleation factor
- PMID: 19287377
- PMCID: PMC2763628
- DOI: 10.1038/ncb1852
p53-cofactor JMY is a multifunctional actin nucleation factor
Abstract
Many cellular structures are assembled from networks of actin filaments, and the architecture of these networks depends on the mechanism by which the filaments are formed. Several classes of proteins are known to assemble new filaments, including the Arp2/3 complex, which creates branched filament networks, and Spire, which creates unbranched filaments. We find that JMY, a vertebrate protein first identified as a transcriptional co-activator of p53, combines these two nucleating activities by both activating Arp2/3 and assembling filaments directly using a Spire-like mechanism. Increased levels of JMY expression enhance motility, whereas loss of JMY slows cell migration. When slowly migrating HL-60 cells are differentiated into highly motile neutrophil-like cells, JMY moves from the nucleus to the cytoplasm and is concentrated at the leading edge. Thus, JMY represents a new class of multifunctional actin assembly factor whose activity is regulated, at least in part, by sequestration in the nucleus.
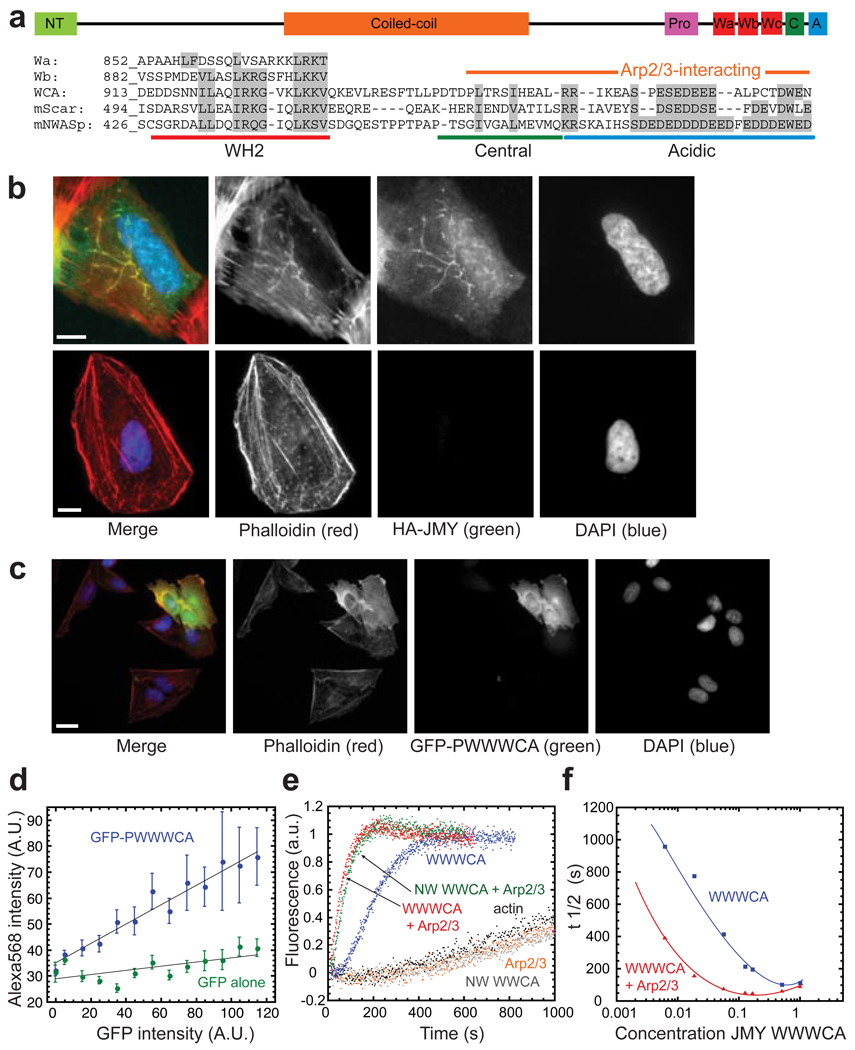
Figures




Comment in
-
Double JMY: making actin fast.Nat Cell Biol. 2009 Apr;11(4):375-6. doi: 10.1038/ncb0409-375. Nat Cell Biol. 2009. PMID: 19337319 Free PMC article.
References
-
- Welch M, Mullins R. Cellular control of actin nucleation. Annu. Rev. Cell Dev. Biol. 2002;18:247–288. - PubMed
-
- Quinlan ME, Heuser JE, Kerkhoff E, Mullins RD. Drosophila Spire is an actin nucleation factor. Nature. 2005;433:382–388. - PubMed
-
- Marchand JB, Kaiser DA, Pollard TD, Higgs HN. Interaction of WASP/Scar proteins with actin and vertebrate Arp2/3 complex. Nat Cell Biol. 2001;3:76–82. - PubMed
-
- Shikama N, et al. A novel cofactor for p300 that regulates the p53 response. Mol. Cell. 1999;4:365–376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
