

The pathway of hepatitis C virus mRNA recruitment to the human ribosome
- PMID: 19287397
- PMCID: PMC2689074
- DOI: 10.1038/nsmb.1572
The pathway of hepatitis C virus mRNA recruitment to the human ribosome
Abstract
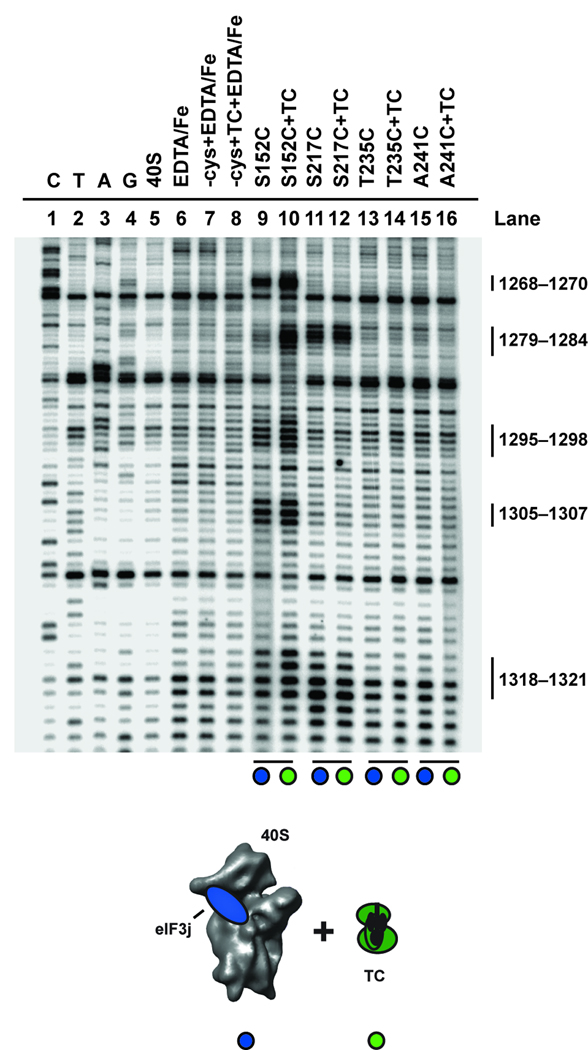
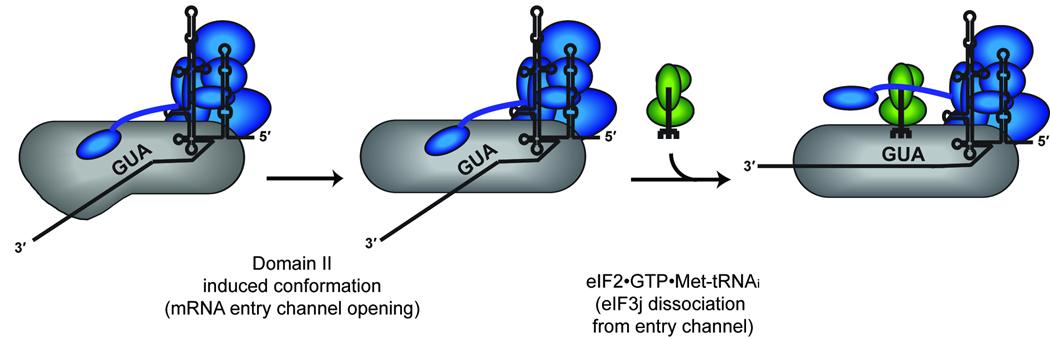
Eukaryotic protein synthesis begins with mRNA positioning in the ribosomal decoding channel in a process typically controlled by translation-initiation factors. Some viruses use an internal ribosome entry site (IRES) in their mRNA to harness ribosomes independently of initiation factors. We show here that a ribosome conformational change that is induced upon hepatitis C viral IRES binding is necessary but not sufficient for correct mRNA positioning. Using directed hydroxyl radical probing to monitor the assembly of IRES-containing translation-initiation complexes, we have defined a crucial step in which mRNA is stabilized upon initiator tRNA binding. Unexpectedly, however, this stabilization occurs independently of the AUG codon, underscoring the importance of initiation factor-mediated interactions that influence the configuration of the decoding channel. These results reveal how an IRES RNA supplants some, but not all, of the functions normally carried out by protein factors during initiation of protein synthesis.
Figures
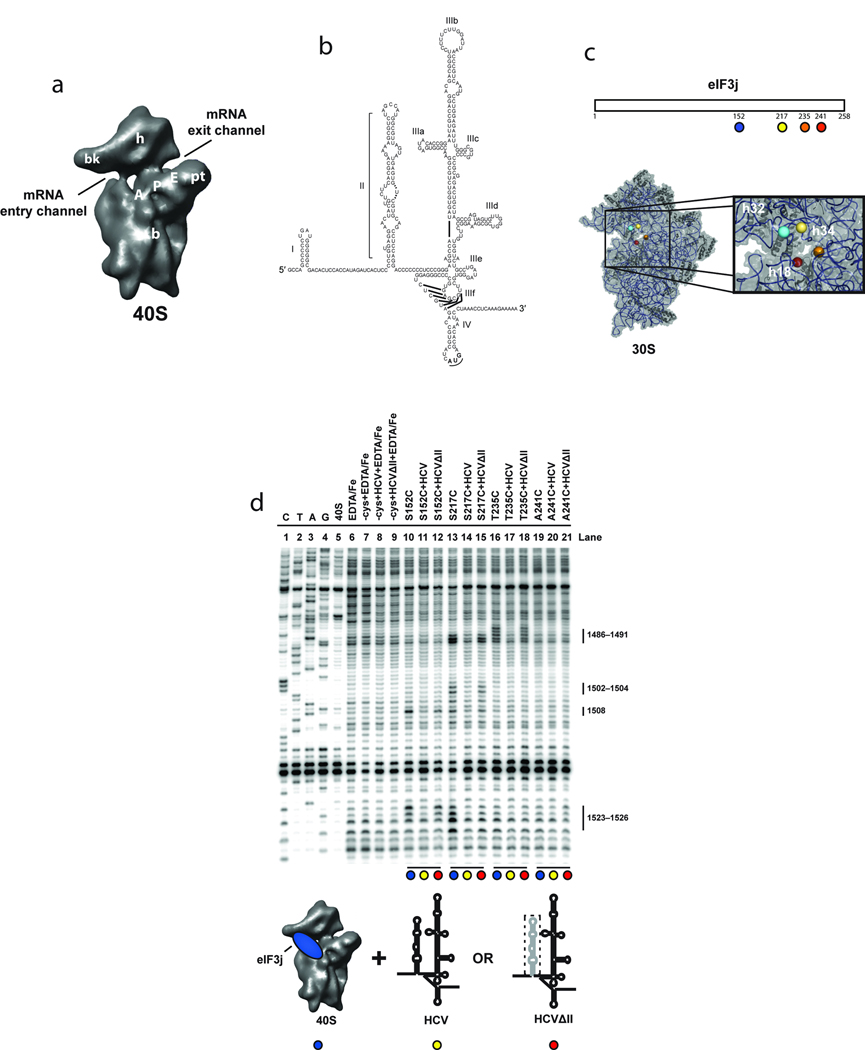
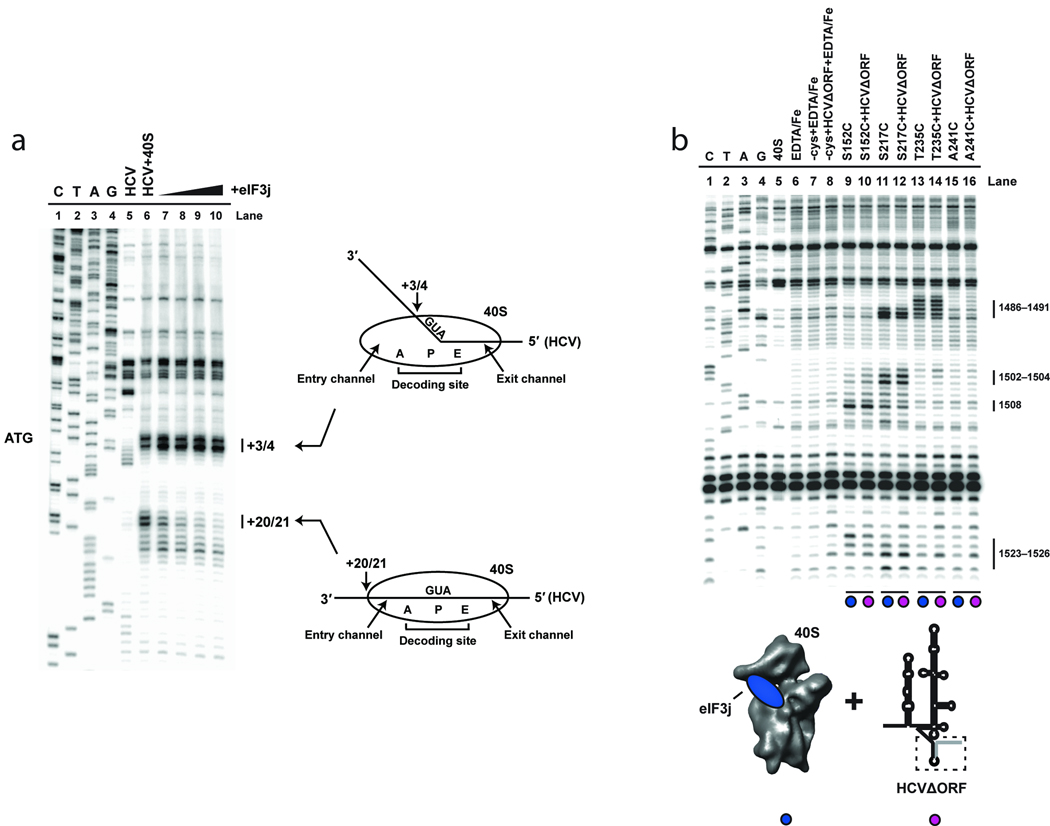
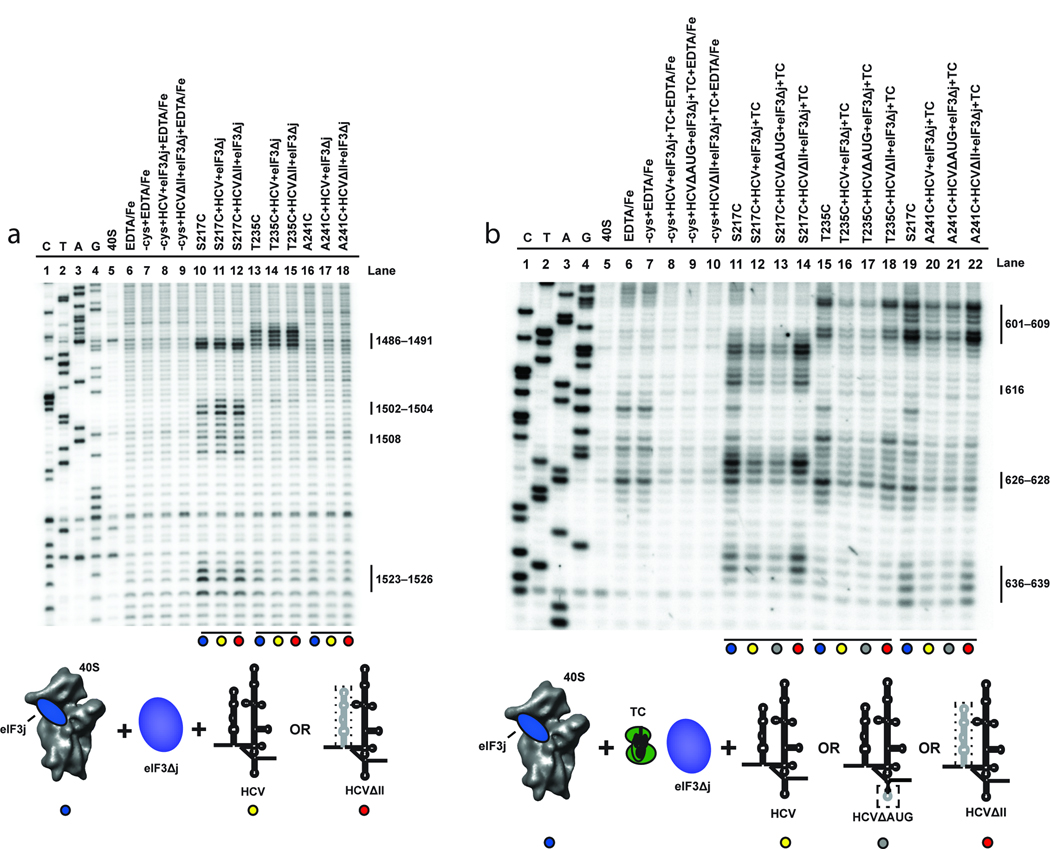
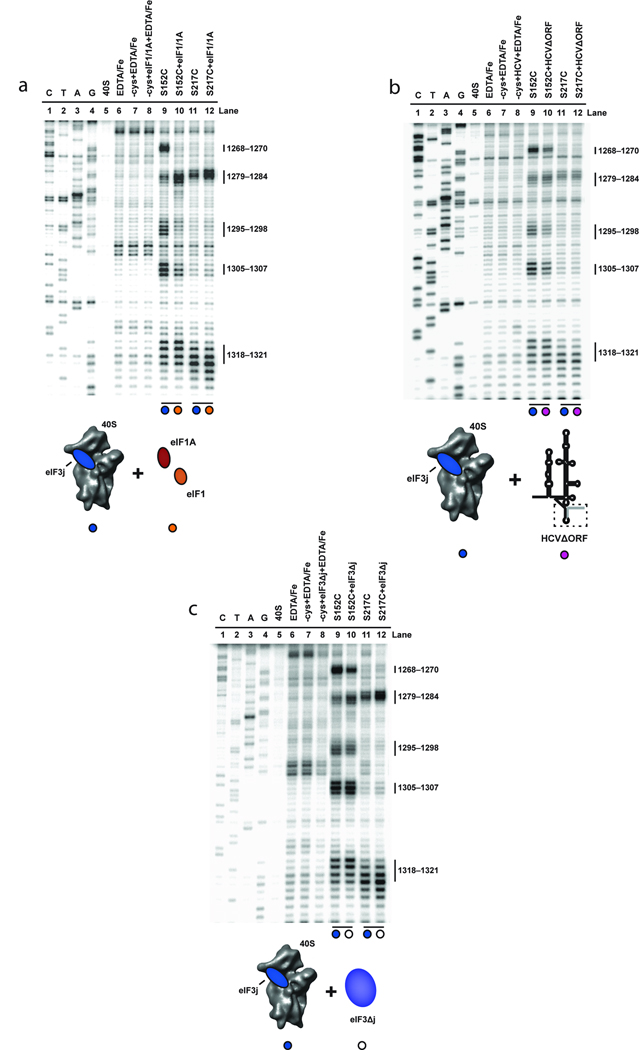
References
-
- Pestova TV, Lorsch JR, Hellen CUT. The mechanism of translation initiation in eukaryotes. In: Mathews MB, Sonenberg N, Hershey JWB, editors. Translational Control in Biology and Medicine. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2007. pp. 87–128.
-
- Fraser CS, Doudna JA. Quantitative studies of ribosome conformational dynamics. Q Rev Biophys. 2007;40:163–189. - PubMed
-
- Doudna JA, Sarnow P. Translation Initiation by Viral Internal Ribosome Entry Sites. In: Mathews MB, Sonenberg N, Hershey JWB, editors. Translational Control in Biology and Medicine. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2007. pp. 129–153.
-
- Elroy-Stein O, Merrick WC. Translation Initiation via Cellular Internal Ribosome Entry Sites. In: Mathews MB, Sonenberg N, Hershey JWB, editors. Translational Control in Biology and Medicine. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2007. pp. 155–172.
-
- Pisarev AV, Shirokikh NE, Hellen CU. Translation initiation by factor-independent binding of eukaryotic ribosomes to internal ribosomal entry sites. C R Biol. 2005;328:589–605. - PubMed
