

Action potential modulates Ca2+-dependent and Ca2+-independent secretion in a sensory neuron
- PMID: 19289069
- PMCID: PMC2907683
- DOI: 10.1016/j.bpj.2008.11.037
Action potential modulates Ca2+-dependent and Ca2+-independent secretion in a sensory neuron
Abstract
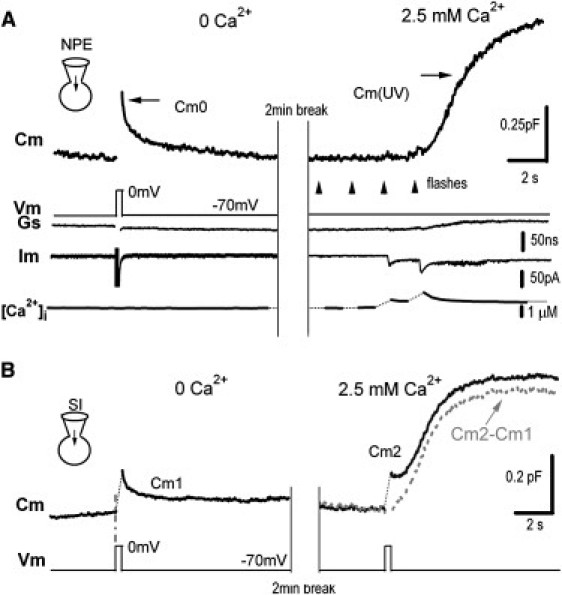
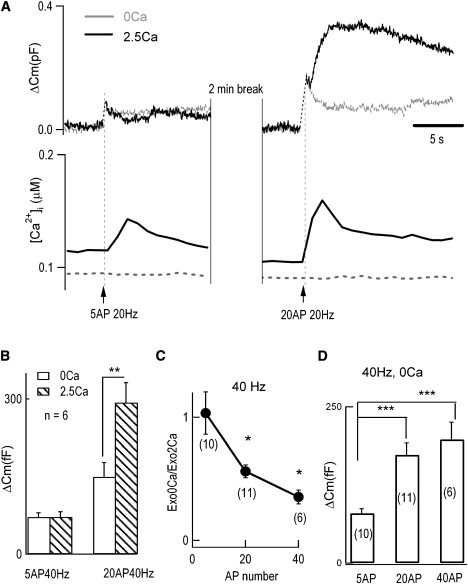
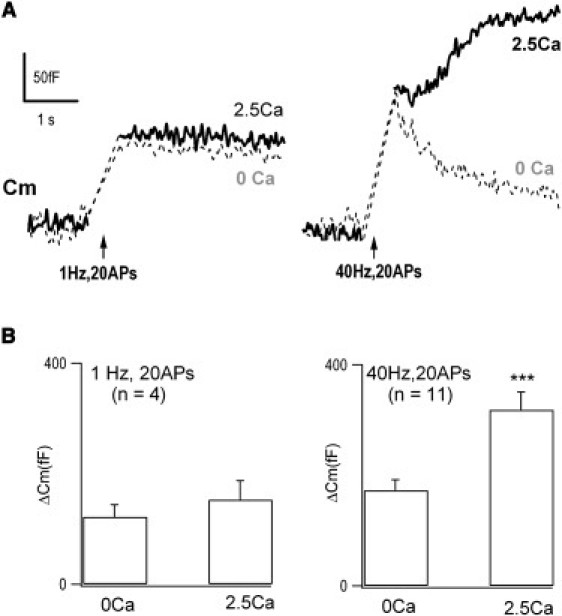
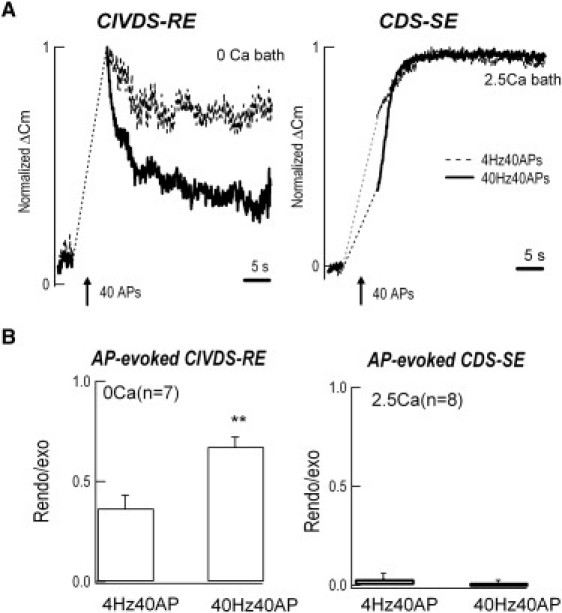
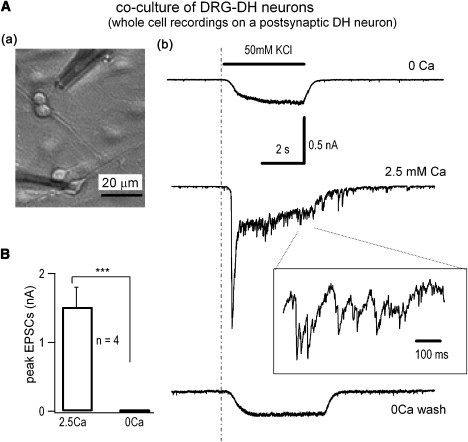
Neurotransmitter release normally requires calcium triggering. However, the somata of dorsal root ganglion (DRG) neurons possess a calcium-independent but voltage-dependent secretion (CIVDS) in addition to the classic calcium-dependent secretion (CDS). Here, we investigated the physiological role of CIVDS and the contributions of CIVDS and CDS induced by action potentials (APs) in DRG soma. Using membrane capacitance measurements, caged calcium photolysis, and membrane capacitance kinetics analysis, we demonstrated that AP-induced secretion had both CIVDS and CDS components. Following physiological stimuli, the dominant component of AP-induced secretion was either CIVDS for spontaneous firing or CDS for high-intensity stimuli. AP frequency modulates CDS-coupled exocytosis and CIVDS-coupled endocytosis but not CIVDS-coupled exocytosis and CDS-coupled endocytosis. Finally, CIVDS did not contribute to excitatory postsynaptic currents induced by APs in DRG presynaptic terminals in the spinal cord. Thus, CIVDS is probably an essential physiological component of AP-induced secretion in the soma. These findings bring novel insights into primary sensory processes in DRG neurons.
Figures
Similar articles
-
Two distinct vesicle pools for depolarization-induced exocytosis in somata of dorsal root ganglion neurons.J Physiol. 2011 Jul 15;589(Pt 14):3507-15. doi: 10.1113/jphysiol.2011.208777. Epub 2011 Jun 6. J Physiol. 2011. PMID: 21646411 Free PMC article.
-
CaV2.2 Gates Calcium-Independent but Voltage-Dependent Secretion in Mammalian Sensory Neurons.Neuron. 2017 Dec 20;96(6):1317-1326.e4. doi: 10.1016/j.neuron.2017.10.028. Epub 2017 Nov 30. Neuron. 2017. PMID: 29198756
-
A simulation study on the Ca2+-independent but voltage-dependent exocytosis and endocytosis in dorsal root ganglion neurons.Eur Biophys J. 2005 Nov;34(8):1007-16. doi: 10.1007/s00249-005-0486-8. Epub 2005 Jun 10. Eur Biophys J. 2005. PMID: 15947975
-
Ca2+ -independent transmission at the central synapse formed between dorsal root ganglion and dorsal horn neurons.EMBO Rep. 2022 Nov 7;23(11):e54507. doi: 10.15252/embr.202154507. Epub 2022 Sep 23. EMBO Rep. 2022. PMID: 36148511 Free PMC article.
-
Calcium- and dynamin-independent endocytosis in dorsal root ganglion neurons.Neuron. 2004 Apr 22;42(2):225-36. doi: 10.1016/s0896-6273(04)00189-8. Neuron. 2004. PMID: 15091339
Cited by
-
An inhibitory effect of extracellular Ca2+ on Ca2+-dependent exocytosis.PLoS One. 2011;6(10):e24573. doi: 10.1371/journal.pone.0024573. Epub 2011 Oct 18. PLoS One. 2011. PMID: 22028769 Free PMC article.
-
Ca2+-independent but voltage-dependent quantal catecholamine secretion (CiVDS) in the mammalian sympathetic nervous system.Proc Natl Acad Sci U S A. 2019 Oct 1;116(40):20201-20209. doi: 10.1073/pnas.1902444116. Epub 2019 Sep 17. Proc Natl Acad Sci U S A. 2019. PMID: 31530723 Free PMC article.
-
Modulation of Kv3.4 channel N-type inactivation by protein kinase C shapes the action potential in dorsal root ganglion neurons.J Physiol. 2012 Jan 1;590(1):145-61. doi: 10.1113/jphysiol.2011.218560. Epub 2011 Nov 7. J Physiol. 2012. PMID: 22063632 Free PMC article.
-
Mechanisms and regulation of neurotrophin synthesis and secretion.Neurosciences (Riyadh). 2016 Oct;21(4):306-313. doi: 10.17712/nsj.2016.4.20160080. Neurosciences (Riyadh). 2016. PMID: 27744458 Free PMC article. Review.
-
Two distinct vesicle pools for depolarization-induced exocytosis in somata of dorsal root ganglion neurons.J Physiol. 2011 Jul 15;589(Pt 14):3507-15. doi: 10.1113/jphysiol.2011.208777. Epub 2011 Jun 6. J Physiol. 2011. PMID: 21646411 Free PMC article.
References
-
- Zucker R.S. Exocytosis: a molecular and physiological perspective. Neuron. 1996;17:1049–1055. - PubMed
-
- Augustine G.J., Charlton M.P., Smith S.J. Calcium action in synaptic transmitter release. Annu. Rev. Neurosci. 1987;10:633–693. - PubMed
-
- Zhou Z., Misler S. Action potential-induced quantal secretion of catecholamines from rat adrenal chromaffin cells. J. Biol. Chem. 1995;270:3498–3505. - PubMed
-
- Neher E., Zucker R.S. Multiple calcium-dependent processes related to secretion in bovine chromaffin cells. Neuron. 1993;10:21–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
