

Fluctuation analysis of tetanic rundown (short-term depression) at a corticothalamic synapse
- PMID: 19289074
- PMCID: PMC2989152
- DOI: 10.1016/j.bpj.2008.12.3891
Fluctuation analysis of tetanic rundown (short-term depression) at a corticothalamic synapse
Abstract
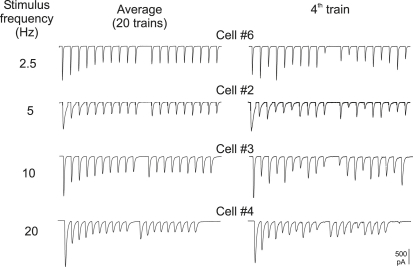
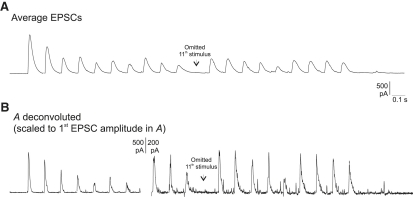
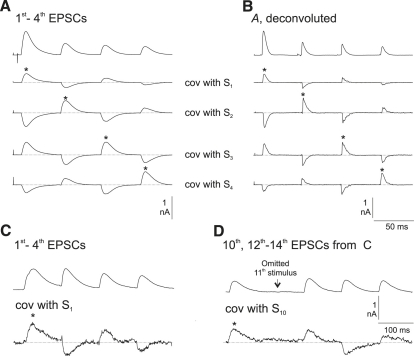
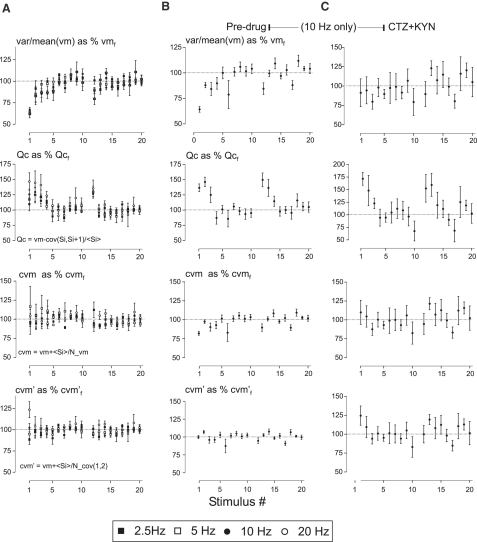
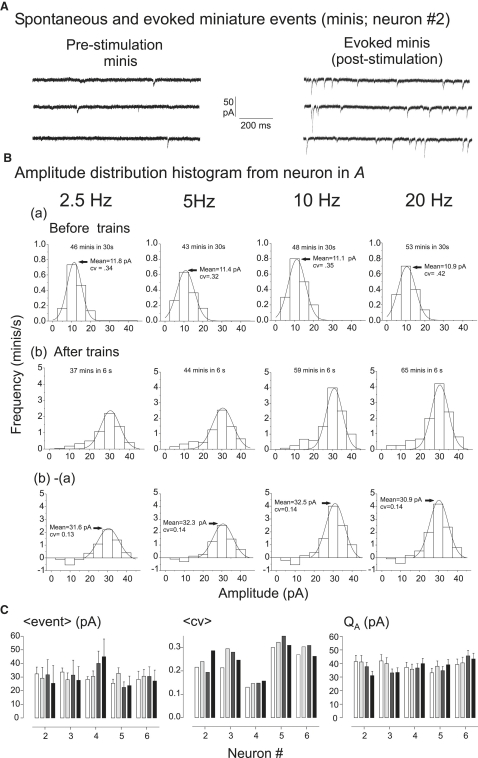
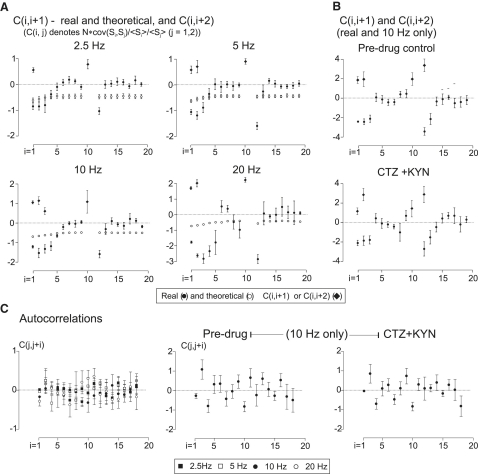
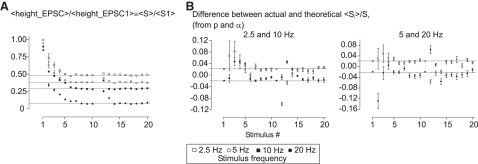
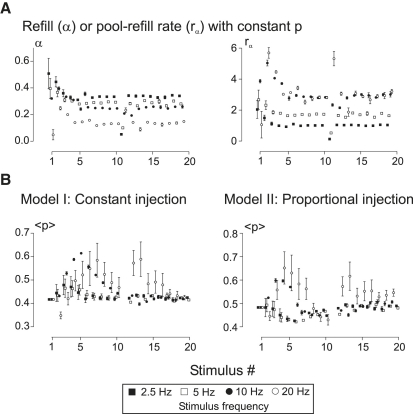
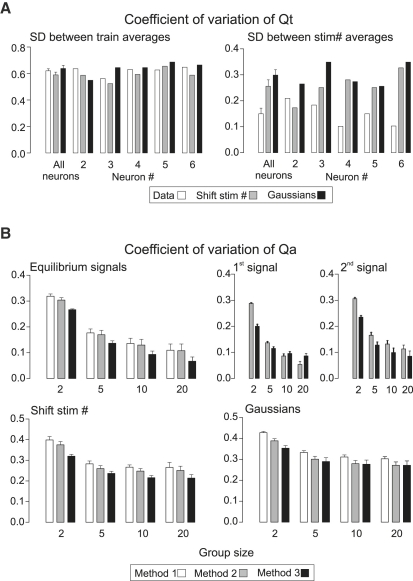
Hypothetical scenarios for "tetanic rundown" ("short-term depression") of synaptic signals evoked by stimulus trains differ in evolution of quantal amplitude (Q) and covariances between signals. With corticothalamic excitatory postsynaptic currents (EPSCs) evoked by 2.5- to 20-Hz trains, we found Q (estimated using various corrections of variance/mean ratios) to be unchanged during rundown and close to the size of stimulus-evoked "miniatures". Except for covariances, results were compatible with a depletion model, according to which incomplete "refill" after probabilistic quantal release entails release-site "emptying". For five neurons with 20 train repetitions at each frequency, there was little between-neuron variation of rundown; pool-refill rate increased with stimulus frequency and evolved during rundown. Covariances did not fit the depletion model or theoretical alternatives, being excessively negative for adjacent EPSCs early in trains, absent at equilibrium, and anomalously positive for some nonadjacent EPSCs. The anomalous covariances were unaltered during pharmacological blockade of receptor desensitization and saturation. These findings suggest that pool-refill rate and release probability at each release site are continually modulated by antecedent outputs in its neighborhood, possibly via feedback mechanisms. In all data sets, sampling errors for between-train variances were much less than theoretical, warranting reconsideration of the probabilistic nature of quantal transmitter release.
Figures


References
-
- Abbott L.F., Varela J.A., Sen K., Nelson S.B. Synaptic depression and cortical gain control. Science. 1997;275:220–224. - PubMed
-
- Chen C., Blitz D.M., Regehr W.G. Contributions of receptor desensitization and saturation to plasticity at the retinogeniculate synapse. Neuron. 2002;33:779–788. - PubMed
-
- Liley A.W., North K.A. An electrical investigation of effects of repetitive stimulation on mammalian neuromuscular junction. J. Neurophysiol. 1953;16:509–527. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
