

Intrinsic tumor suppression and epithelial maintenance by endocytic activation of Eiger/TNF signaling in Drosophila
- PMID: 19289090
- PMCID: PMC2729686
- DOI: 10.1016/j.devcel.2009.01.002
Intrinsic tumor suppression and epithelial maintenance by endocytic activation of Eiger/TNF signaling in Drosophila
Abstract
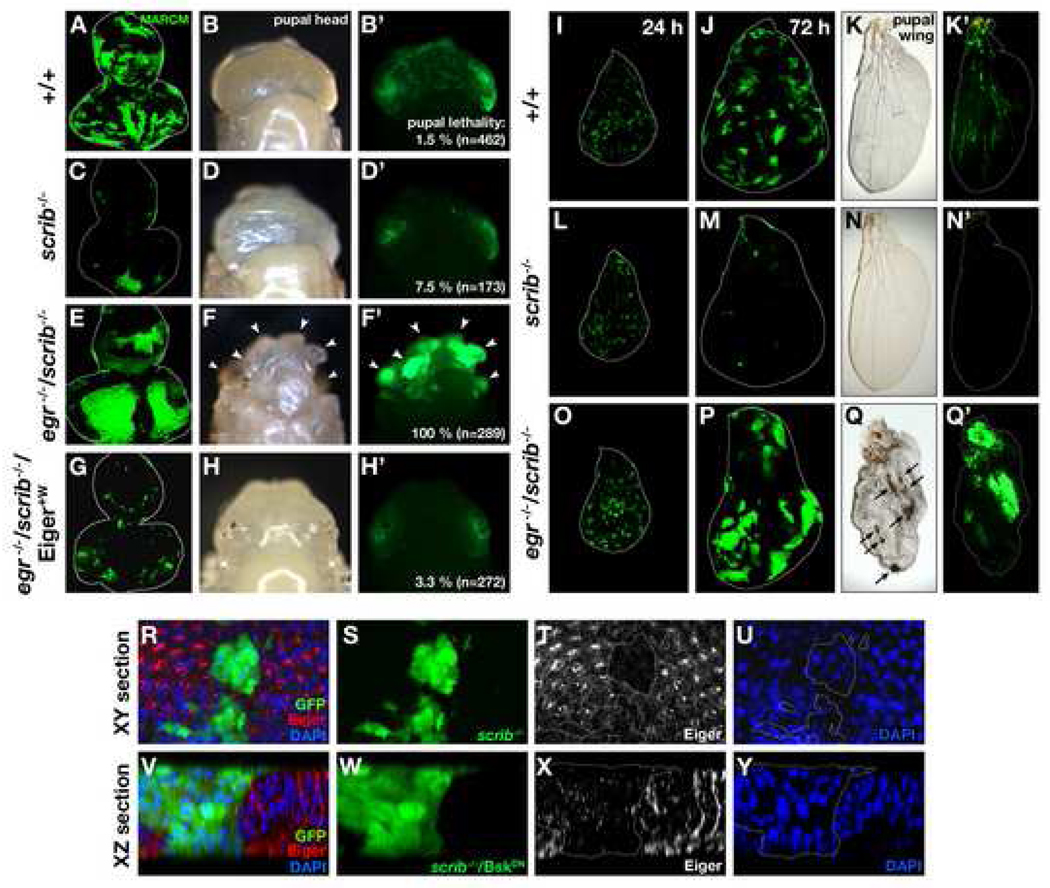
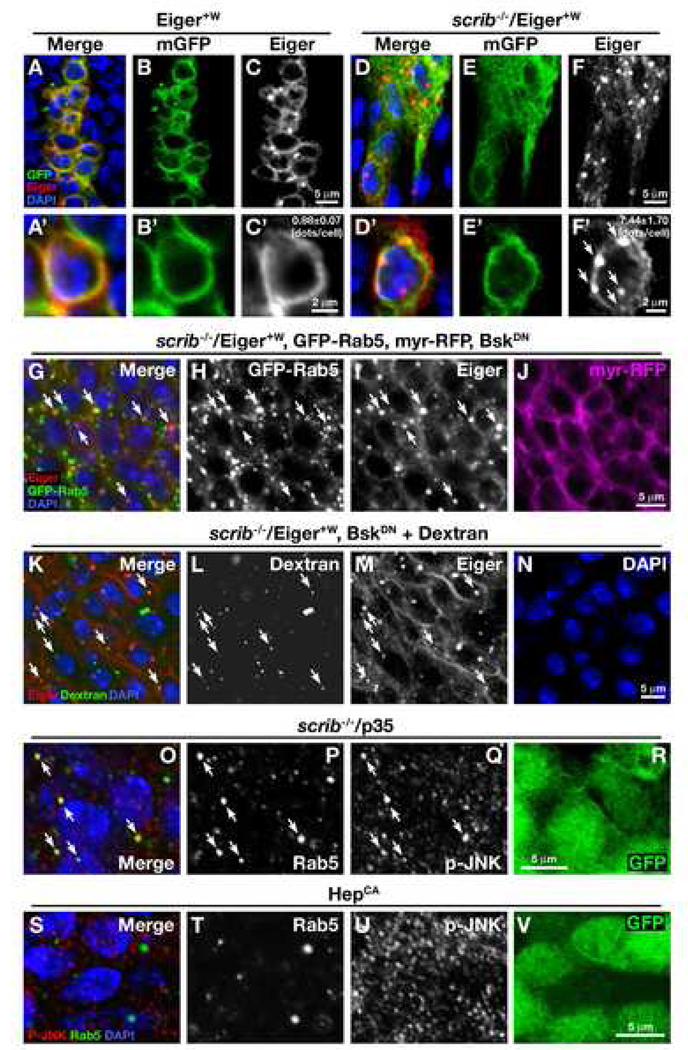
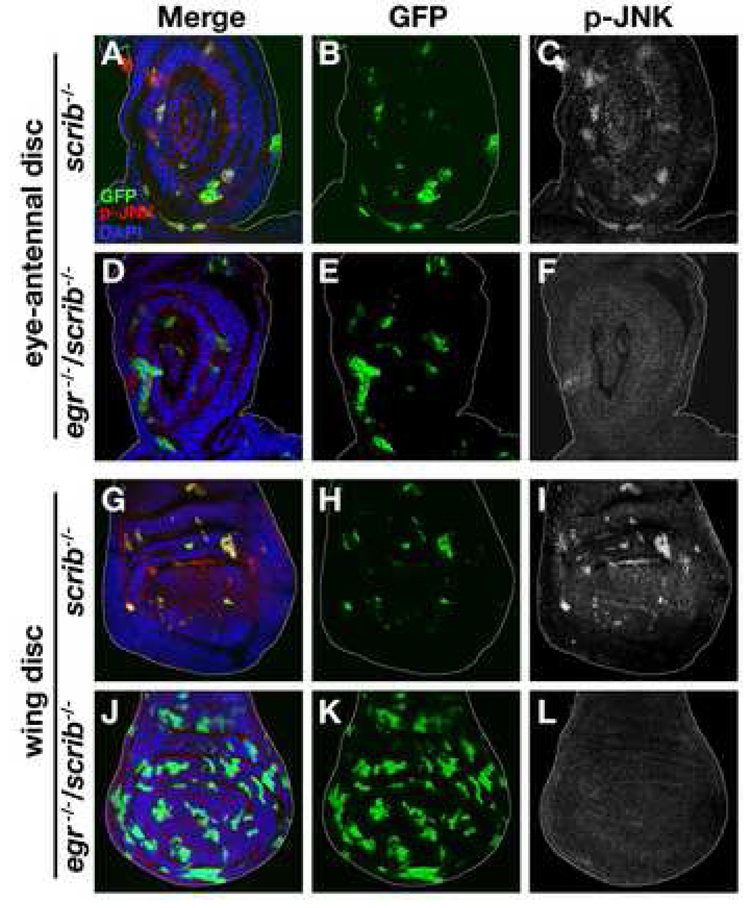
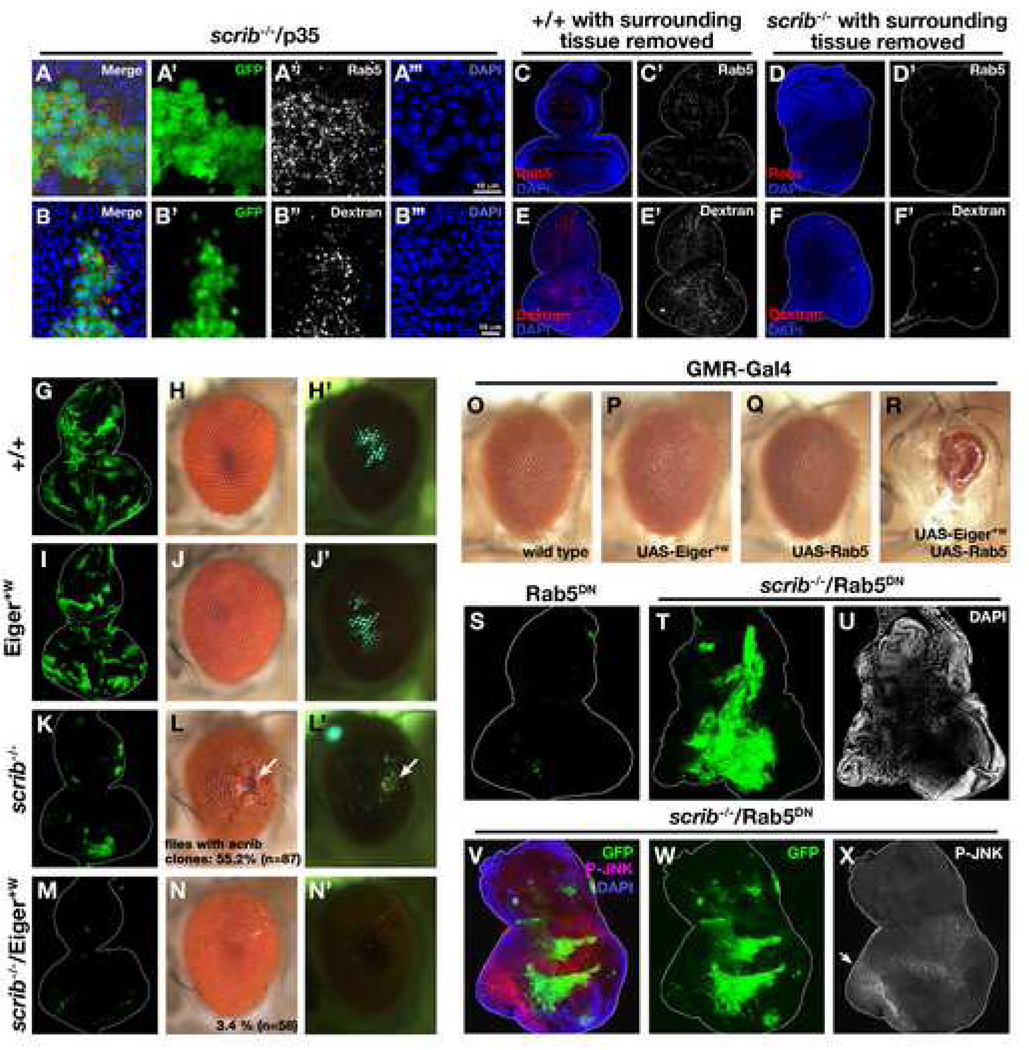
Oncogenic alterations in epithelial tissues often trigger apoptosis, suggesting an evolutionary mechanism by which organisms eliminate aberrant cells from epithelia. In Drosophila imaginal epithelia, clones of cells mutant for tumor suppressors, such as scrib or dlg, lose their polarity and are eliminated by cell death. Here, we show that Eiger, the Drosophila tumor necrosis factor (TNF), behaves like a tumor suppressor that eliminates oncogenic cells from epithelia through a local endocytic JNK-activation mechanism. In the absence of Eiger, these polarity-deficient clones are no longer eliminated; instead, they grow aggressively into tumors. We show that in scrib clones endocytosis is elevated, which translocates Eiger to endocytic vesicles and leads to activation of apoptotic JNK signaling. Furthermore, blocking endocytosis prevents both JNK activation and cell elimination. Our data indicate that TNF signaling and the endocytic machinery could be components of an evolutionarily conserved fail-safe mechanism by which animals protect against neoplastic development.
Figures
References
-
- Adachi-Yamada T, Fujimura-Kamada K, Nishida Y, Matsumoto K. Distortion of proximodistal information causes JNK-dependent apoptosis in Drosophila wing. Nature. 1999a;400:166–169. - PubMed
-
- Aggarwal BB. Signalling pathways of the TNF superfamily: a double-edged sword. Nat Rev Immunol. 2003;3:745–756. - PubMed
-
- Agnes F, Suzanne M, Noselli S. The Drosophila JNK pathway controls the morphogenesis of imaginal discs during metamorphosis. Development. 1999;126:5453–5462. - PubMed
-
- Agrawal N, Kango M, Mishra A, Sinha P. Neoplastic transformation and aberrant cell-cell interactions in genetic mosaics of lethal(2)giant larvae (lgl), a tumor suppressor gene of Drosophila. Dev Biol. 1995;172:218–229. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
