

Yersinia pestis can reside in autophagosomes and avoid xenophagy in murine macrophages by preventing vacuole acidification
- PMID: 19289509
- PMCID: PMC2687347
- DOI: 10.1128/IAI.00068-09
Yersinia pestis can reside in autophagosomes and avoid xenophagy in murine macrophages by preventing vacuole acidification
Abstract
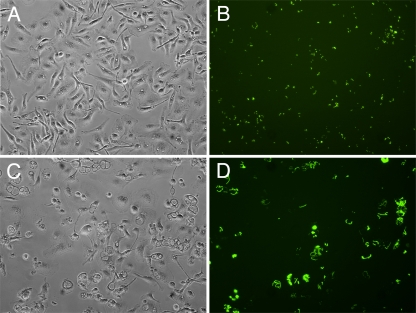
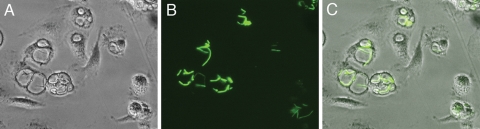
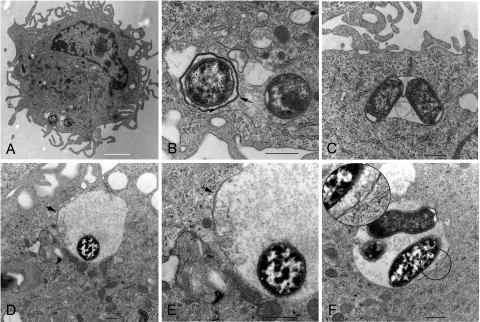
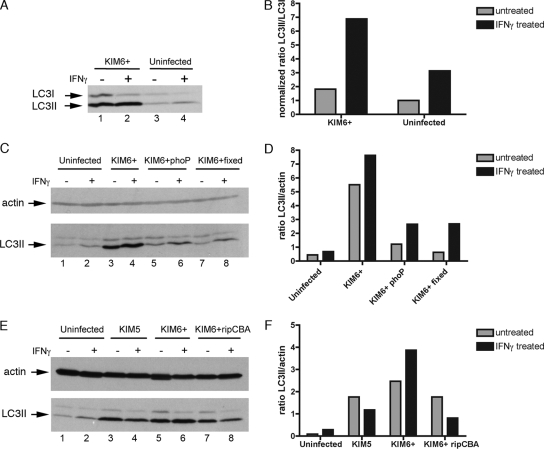
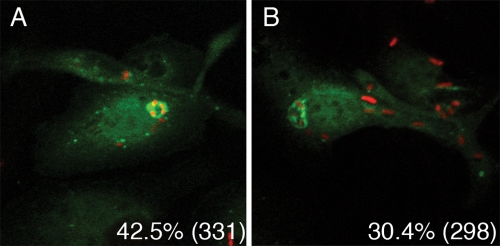
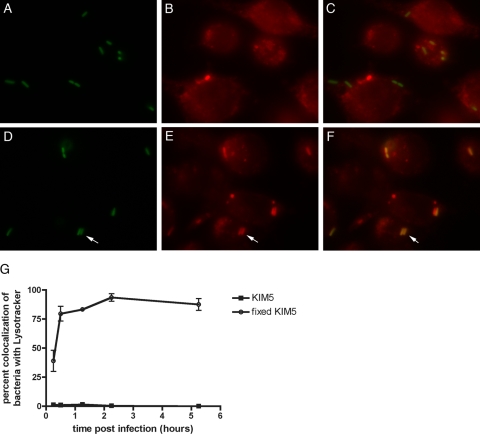
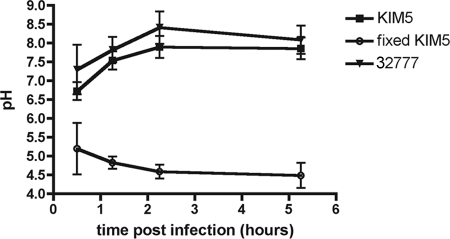
Yersinia pestis survives and replicates in phagosomes of murine macrophages. Previous studies demonstrated that Y. pestis-containing vacuoles (YCVs) acquire markers of late endosomes or lysosomes in naïve macrophages and that this bacterium can survive in macrophages activated with the cytokine gamma interferon. An autophagic process known as xenophagy, which destroys pathogens in acidic autophagolysosomes, can occur in naïve macrophages and is upregulated in activated macrophages. Studies were undertaken here to investigate the mechanism of Y. pestis survival in phagosomes of naïve and activated macrophages and to determine if the pathogen avoids or co-opts autophagy. Colocalization of the YCV with markers of autophagosomes or acidic lysosomes and the pH of the YCV were determined by microscopic imaging of infected macrophages. Some YCVs contained double membranes characteristic of autophagosomes, as determined by electron microscopy. Fluorescence microscopy showed that approximately 40% of YCVs colocalized with green fluorescent protein (GFP)-LC3, a marker of autophagic membranes, and that YCVs failed to acidify below pH 7 in naïve macrophages. Replication of Y. pestis in naïve macrophages caused accumulation of LC3-II, as determined by immunoblotting. While activation of infected macrophages increased LC3-II accumulation, it decreased the percentage of GFP-LC3-positive YCVs (approximately 30%). A viable count assay showed that Y. pestis survived equally well in macrophages proficient for autophagy and macrophages rendered deficient for this process by Cre-mediated deletion of ATG5, revealing that this pathogen does not require autophagy for intracellular replication. We conclude that although YCVs can acquire an autophagic membrane and accumulate LC3-II, the pathogen avoids xenophagy by preventing vacuole acidification.
Figures
References
-
- Achtman, M., G. Morelli, P. Zhu, T. Wirth, I. Diehl, B. Kusecek, A. J. Vogler, D. M. Wagner, C. J. Allender, W. R. Easterday, V. Chenal-Francisque, P. Worsham, N. R. Thomson, J. Parkhill, L. E. Lindler, E. Carniel, and P. Keim. 2004. Microevolution and history of the plague bacillus, Yersinia pestis. Proc. Natl. Acad. Sci. USA 10117837-17842. - PMC - PubMed
-
- Alexeyev, M. F. 1995. Three kanamycin resistance gene cassettes with different polylinkers. BioTechniques 1852, 54, 56. - PubMed
-
- Amano, A., I. Nakagawa, and T. Yoshimori. 2006. Autophagy in innate immunity against intracellular bacteria. J. Biochem. 140161-166. - PubMed
-
- Birmingham, C. L., V. Canadien, N. A. Kaniuk, B. E. Steinberg, D. E. Higgins, and J. H. Brumell. 2008. Listeriolysin O allows Listeria monocytogenes replication in macrophage vacuoles. Nature 451350-354. - PubMed
-
- Cavanaugh, D. C., and R. Randall. 1959. The role of multiplication of Pasteurella pestis in mononuclear phagocytes in the pathogenesis of fleaborne plague. J. Immunol. 85348-363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
