

Sensitization of lamina I spinoparabrachial neurons parallels heat hyperalgesia in the chronic constriction injury model of neuropathic pain
- PMID: 19289544
- PMCID: PMC2689339
- DOI: 10.1113/jphysiol.2009.170290
Sensitization of lamina I spinoparabrachial neurons parallels heat hyperalgesia in the chronic constriction injury model of neuropathic pain
Abstract
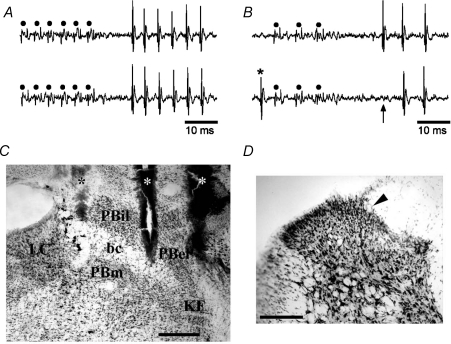
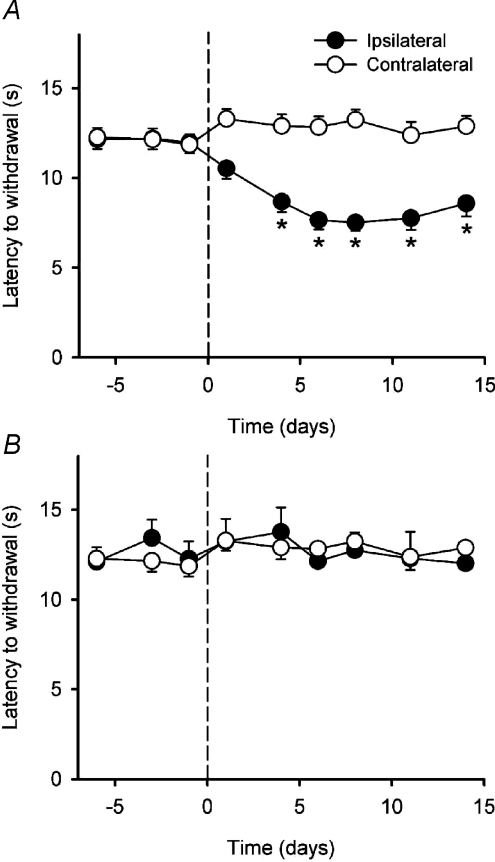
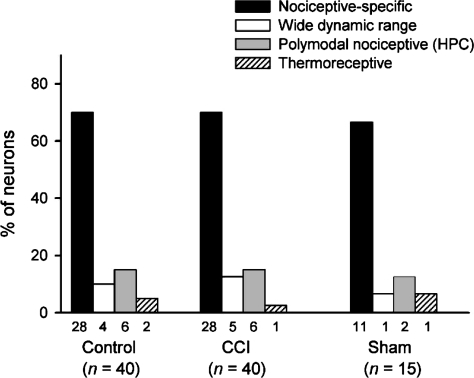
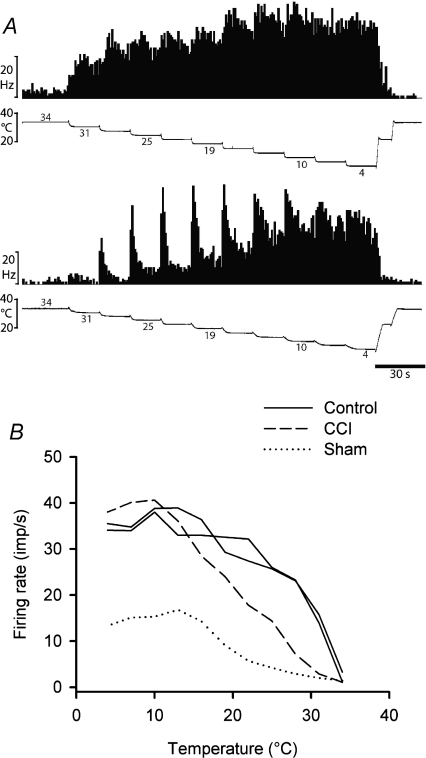
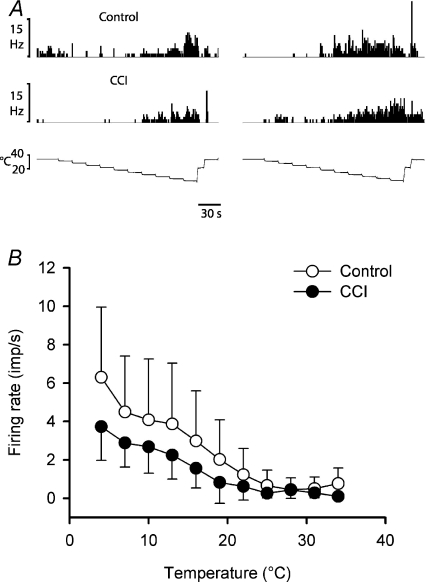
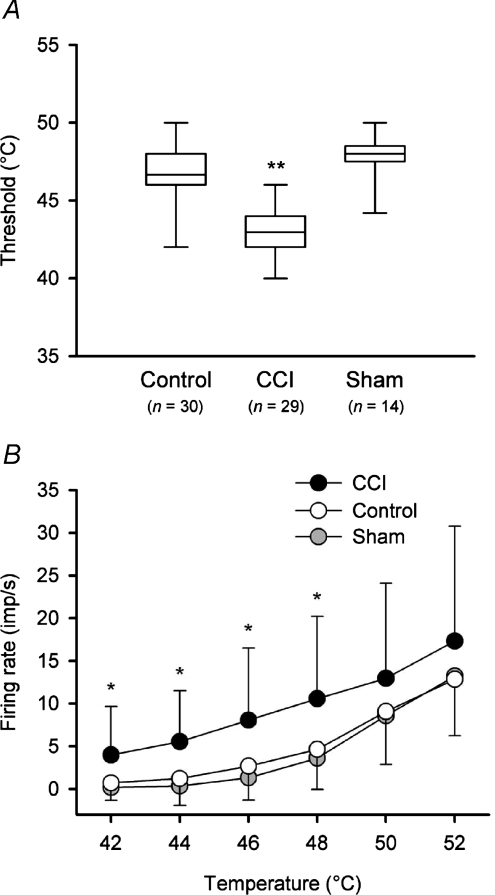
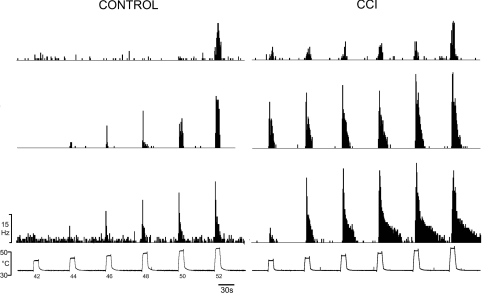
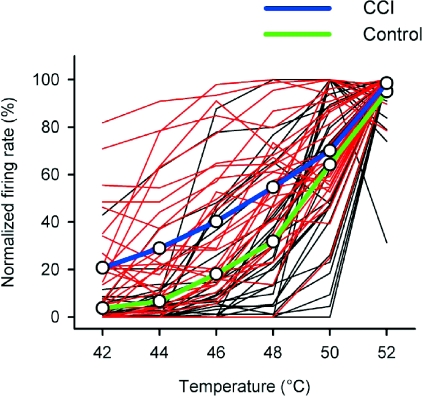
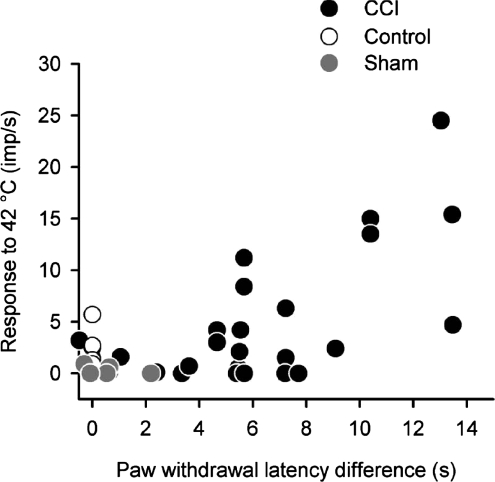
It has been proposed that spinal lamina I neurons with ascending axons that project to the midbrain play a crucial role in hyperalgesia. To test this hypothesis the quantitative properties of lamina I spinoparabrachial neurons in the chronic constriction injury (CCI) model of neuropathic pain were compared to those of unoperated and sham-operated controls. Behavioural testing showed that animals with a CCI exhibited heat hyperalgesia within 4 days of the injury, and this hyperalgesia persisted throughout the 14-day post-operative testing period. In the CCI, nociceptive lamina I spinoparabrachial neurons had heat thresholds that were significantly lower than controls (43.0 +/- 2.8 degrees C vs. 46.7 +/- 2.6 degrees C; P < 10(-4), ANOVA). Nociceptive lamina I spinoparabrachial neurons were also significantly more responsive to graded heat stimuli in the CCI, compared to controls (P < 0.02, 2-factor repeated-measures ANOVA), and increased after-discharges were also observed. Furthermore, the heat-evoked stimulus-response functions of lamina I spinoparabrachial neurons in CCI animals co-varied significantly (P < 0.03, ANCOVA) with the amplitude of heat hyperalgesia determined behaviourally. Taken together these results are consistent with the hypothesis that lamina I spinoparabrachial neurons have an important mechanistic role in the pathophysiology of neuropathic pain.
Figures
References
-
- Ahlgren SC, White DM, Levine JD. Increased responsiveness of sensory neurons in the saphenous nerve of the streptozotocin-diabetic rat. J Neurophysiol. 1992;68:2077–2085. - PubMed
-
- Attal N, Jazat F, Kayser V, Guilbaud G. Further evidence for ‘pain-related’ behaviours in a model of unilateral peripheral neuropathy. Pain. 1990;41:235–251. - PubMed
-
- Balasubramanyan S, Stemkowski PL, Stebbing MJ, Smith PA. Sciatic chronic constriction produces cell-type-specific changes in the electrophysiological properties of rat substantia gelatinosa neurons. J Neurophysiol. 2006;96:579–590. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources